Comportamientos Relacionados con la Justicia y la Equidad en Primates No Humanos
Sarah F. Brosnan
ABSTRACT. A distinctive feature across human societies is our interest in justice and fairness. People will sometimes invest in extremely costly behavior to achieve fair outcomes for themselves and others. Why do people care so much about justice? One way to address this is comparatively, exploring behaviors related to justice and fairness in other species. In this paper, I review work exploring responses to inequity, prosocial behavior, and other relevant behaviors in nonhuman primates in an effort to understand both the potential evolutionary function of these behaviors and the social and ecological reasons for the individual differences in behavior. I also consider how these behaviors relate to human behavior, particularly in the case of experimental studies using games derived from experimental economics to compare nonhuman primates’ responses to those of humans in similar experimental conditions. These results emphasize the importance of a comparative approach to better understand the function and diversity of human behavior.
KEYWORDS. Comparative economics, decision-making, game theory, behavioral phylogeny, cooperation.
Un rasgo humano que al parecer nos separa como especie es nuestro interés en el bienestar de los demás y de nosotros mismos (Eisenberg y Mussen 1989; Henrich, et al. 2004). En ocasiones, a pesar del costo personal que implica, protestamos ante resultados que consideramos inequitativos, sin importar si nos afectan personalmente o aunque no tengan un impacto directo en nuestras vidas. ¿Qué es lo que hace que la gente se comporte de esta manera? Una posibilidad es que se puede atribuir a la cultura. De hecho, solamente en inglés existe el concepto de fairness (que aquí se ha traducido como ‘equidad’); otras lenguas han adoptado la palabra inglesa, lo que indica un amplio interés cultural en el concepto, aunque al mismo tiempo, implica que el concepto no es ubicuo (Wilson 2012). Otra posibilidad es que hayamos evolucionado para interesarnos en el bienestar de los demás. Aunque hay claros beneficios vinculados con la eficacia biológica por la consideración del bienestar de uno mismo, hay también beneficios (aunque en un plazo mayor) por la consideración del bienestar de los demás. Puede haber costos por ayudar a otro en el corto plazo, pero si también hay beneficios a largo plazo, ya sea a causa de interacciones continuas basadas en la reciprocidad, o de manera más indirecta, a causa del incremento en la reputación como una persona “equitativa”, entonces la conducta puede ser seleccionada (Frank 1988). Si esta conducta ha evolucionado deberíamos encontrar conductas similares, o precursores conductuales, en otras especies. Las otras especies no mostrarán esta conducta de la misma manera que se observa en los humanos. Entre otras razones porque conductas relacionadas con la equidad y la justicia, tales como la capacidad para compartir información sobre la conducta previa de terceros, requieren lenguaje. No obstante, una pregunta importante es el grado al cual la justicia y la equidad han evolucionado de formas similares de conductas en otras especies. Este abordaje evolucionista provee una revelación en la comprensión de nuestra propia conducta.
En los humanos, el concepto de equidad está estrechamente relacionado con el de justicia. Para distinguirlos, la equidad involucra interacciones voluntarias con otros individuos, mientras que la justicia se impone por un tercer partido imparcial (Wilson 2012). Con base en estas definiciones, la mayor parte del trabajo experimental que se discute aquí se relaciona con la equidad, más que con la justicia, ya que los sujetos que se han estudiado se involucran en interacciones y resultados en pugna. Fuera del contexto experimental, las observaciones sugieren que hay intervenciones imparciales en las interacciones con los demás, lo cual indica que estas especies tienen cierta sensibilidad a la justicia. Hay diversos aspectos de la justicia y la equidad, donde se incluyen la distribución, lo procesal, el castigo, la restauración, y demás. La mayor parte de los estudios experimentales con especies no humanas miden la respuesta de los sujetos en diferentes distribuciones de recompensas alimentarias y, por lo tanto, tienen que ver con la justicia distributiva y la equidad. En los humanos, dos mecanismos comunes de la distribución son el igualitarismo, en el cual todos los individuos reciben el mismo resultado (p.ej., igualdad), y la equidad, en la cual los individuos reciben beneficios proporcionales a su inversión. Ambos son adecuados para ser estudiados en otras especies y son el rasgo más comúnmente manipulado en los ya mencionados estudios experimentales (p.ej., por medio de la alteración de la recompensa comparada con la de sus compañeros). Fuera del mundo experimental, los estudios de observación indican que los primates no humanos también son sensibles a los aspectos no distributivos de la justicia y la equidad, tales como el juicio sobre la pertinencia de la conducta de otro individuo, lo cual provee un contexto más amplio en donde considerar la justicia y equidad en estas especies.
El estudio de la conducta de otras especies en estas situaciones nos permite elucidar la función evolutiva de la justicia y la equidad por medio de la exploración de conductas relacionadas, ya sea porque la conducta en sí misma es un componente de la justicia y la equidad o porque ésta representa un precursor de la conducta humana. El estudio de otras especies nos da información en dos caminos complementarios, a través de la homología y la convergencia. La homología sucede cuando una conducta aparece en múltiples especies en tanto que esa conducta también estaba presente en el ancestro común. Esto señala cuándo en la historia filogenética del taxón el rasgo evolucionó. La convergencia se da cuando una conducta evoluciona en múltiples especies como consecuencia de presiones selectivas comunes pero sin ascendencia compartida de un ancestro común. Las convergencias pueden en este caso ser particularmente informativas cuando se estudia la función de una conducta ya que por medio de la comparación entre especies, pueden indicarnos qué rasgos se comparten y las presiones sociales y ecológicas que pudieron ser seleccionadas para esa conducta.
Mi interés en esta revisión es reunir los datos con respecto a las conductas que se relacionan con la justicia y la equidad en especies de primates no humanos (a partir de aquí, primates). Algunas observaciones han destacado las situaciones en que los primates actúan como si su conducta estuviera guiada por el interés en los resultados de los demás (de Waal 1991). Hace una década, los primeros estudios experimentales surgieron y se enfocaron en un aspecto de la justicia y la equidad: cómo respondían los sujetos cuando recibían menos que un compañero (Brosnan y de Waal 2003; Brosnan, et al. 2005). Estos estudios utilizaron métodos derivados de la teoría de juegos y mostraron que, en condiciones experimentales en los que la inequidad entre dos individuos fue introducida, los sujetos respondieron negativamente a la inequidad cuando ésta se presentaba en su contra. A partir de ese momento, esta tarea se ha extendido a catorce especies, que incluyen diez especies de primates, lo cual permite una amplia visión filogenética que proporciona una mirada a la función evolutiva de la inequidad (Brosnan 2011). Además, las respuestas a la inequidad han sido exploradas en contextos y situaciones específicas en cada especie (Brosnan, et al. 2006; de Waal y Berger 2000). Los resultados destacan la correlación entre la inequidad y la cooperación, al mismo tiempo que enfatizan los factores socioecológicos que influenciaron la emergencia de la conducta relacionada con la justicia y la equidad. Estos análisis comparativos comienzan a dar luz sobre los factores que podrían haber llevado a la evolución de conductas relacionadas con la justicia y la equidad en humanos y otros primates.
CONDUCTAS RELACIONADAS CON LA JUSTICIA Y LA EQUIDAD EN OTRAS ESPECIES
Una hipótesis para la evolución de la equidad es que el reconocimiento de inequidades ayudó a los individuos a cerciorase del valor de sus compañeros en interacciones cooperativas (Brosnan 2011; Fehr y Schmidt 1999). Los individuos no deben continuar trabajando con otros si ellos no se benefician, en la misma medida, por dicha relación. Si los resultados son equitativos deben utilizarse como un valor representativo; si los resultados de uno son deseados por el otro de manera rutinaria, entonces es probable que una interacción con otro compañero, incluso uno elegido al azar, lleve a un resultado más benéfico. Esto conduce a un mecanismo de elección de compañero en donde los individuos prueban a otros compañeros cuando ellos experimentan altos niveles de inequidad en la interacción con los compañeros o compañero en curso (Brosnan 2006). Nótese que los individuos, por lo tanto, reconocen y responden a la inequidad sin comprender por qué lo están haciendo. Además, esto proporciona un beneficio incluso cuando los resultados de los individuos en la interacción menos ventajosa son positivamente netos, ya que el aporte de la selección natural es una ganancia relativa y no absoluta.
Aunque esta hipótesis que vincula la inequidad con la cooperación emergió en el contexto humano, otras especies también cooperan entre sí, así que las conductas relacionadas con la inequidad pueden ocurrir en otras especies cooperativas. El abordaje comparativo es complicado. Primero, las manifestaciones de justicia y equidad seguramente son diferentes en distintas especies, lo cual hace difícil para los experimentadores humanos el reconocimiento de conductas relevantes. Segundo, las especies no humanas son también no verbales, con la doble implicación de que no podemos preguntarles sobre la motivación de sus conductas, como tampoco podemos preguntarles sobre los cambios que no se expresan de forma conductual. Por lo tanto, perderemos cualquier respuesta que no tenga una manifestación conductual. En cuanto a resultados, los experimentos se concentran en la forma en que una especie actúa y no en la manera en que los individuos de esa especie quieren actuar. Finalmente, la limitante en la medición de los cambios conductuales significa que los métodos que usualmente se utilizan en los humanos difieren sustancialmente de aquellos utilizados en otros primates, lo cual puede complicar el análisis comparativo.
Hay dos abordajes en el trabajo comparativo relacionado con la justicia y la equidad. El primero es adaptar los métodos de la teoría de juegos para explorar la toma de decisiones en otras especies. La teoría de juegos es una aproximación robusta que permite modelar la compleja toma de decisiones de forma experimental a partir de una serie de opciones cuidadosamente controladas, que pueden ser experimentalmente manipuladas para explorar rigurosamente la conducta que interesa (Smith 1987). La ventaja de la teoría de juegos para la investigación comparativa es que se concentra en opciones simples que permiten desentrañar el comportamiento complejo, lo que da lugar a la posibilidad de utilizar estas técnicas en una amplia variedad de especies. Hay procedimientos que se diseñan que no requieren instrucciones verbales de ningún tipo y que pueden emplearse de la misma manera en humanos y en otras especies, lo que permite comparaciones directas y análisis subsecuentes de resultados o mecanismos compartidos (Brosnan, et al., en dictamen). Aun cuando no podemos asumir que diferentes especies (o diferentes individuos dentro de una misma especie) interpretan los mismos procedimientos de la misma manera, si dichos procedimientos difieren entre especies, entonces nunca seremos capaces de desentrañar si las diferencias resultantes en una conducta se pueden atribuir a las diferencias específicas o a las diferencias en el procedimiento. Una de las complicaciones de las aproximaciones experimentales es que los humanos interactúan con un experimentador de su misma especie, mientras que otras especies realizan dicha interacción con un experimentador de otra especie (un humano). Aunque esto es una necesidad práctica, algunas especies no evolucionaron para interactuar con los humanos, por lo cual se necesitan controles (experimentales) muy cuidadosos para determinar si los sujetos no humanos evalúan sus resultados de manera relativa a sus compañeros conespecíficos, y no al experimentador o a otros aspectos del ambiente. Además, las aproximaciones experimentales son artificiales, y por ello es posible que perdamos de vista algunas situaciones en que la conducta relacionada con la justicia ocurre. A pesar de todo, las aproximaciones de la teoría de juegos tienen la ventaja de ser comparables con la experiencia humana y por eso son indispensables para la investigación comparativa.
La segunda aproximación es la exploración de conductas que pueden relacionarse con la justicia y la equidad en contextos específicos para cada especie. Aquí se incluyen los estudios de observación en los que la conducta es observada en interacciones no controladas. Este abordaje tiene la ventaja de ser el más natural, lo que conduce a probables conductas distinguibles y a un mejor entendimiento de cómo la justicia y la equidad se manifiestan en las interacciones típicas de la especie. Por otro lado, dado que estos estudios no requieren la manipulación de resultados y la medición de las conductas subsecuentes del sujeto, existe la oportunidad de estudiar la justicia fuera del contexto de la recompensa alimenticia. La desventaja es que los resultados no son comparables entre especies, particularmente si los métodos o los contextos en los que los individuos atienden a la inequidad difieren y las motivaciones conductuales subyacentes de los sujetos no se conocen. Lo ideal es combinar elementos de ambos enfoques.
LA UTILIZACIÓN DE LA TEORÍA DE JUEGOS PARA EXPLORAR CONDUCTAS RELACIONADAS CON LA JUSTICIA
El Juego del Ultimátum
En 1982, Werner Guth presentó el Juego del Ultimátum (JU), diseñado para explorar la equidad de las negociaciones entre humanos (Guth, et al. 1982) (Fig. 1). En este juego, un individuo, el que propone, se le da una cantidad fija y se le dice que lo distribuya entre su compañero o compañera y él mismo. Si el compañero, al que se le denomina ‘el que responde’, acepta la distribución, ambas partes reciben su dinero tal como se repartió, pero si el que responde rechaza la distribución, ninguna de las dos partes recibe nada. Cientos de experimentos en las últimas tres décadas han mostrado que los humanos modernos occidentales distribuyen regularmente alrededor del 40 por ciento de la suma total para el que responde, y los que responden regularmente rechazan cualquier oferta menor al 20 por ciento de la distribución total (Camerer 2003). Las distribuciones son mayores que aquellas observadas en un juego similar, el Juego del Dictador (JD), en el que el que responde no tiene manera de rechazar la distribución del que propone, lo cual indica que el que propone altera sus distribuciones dependiendo de la capacidad del que responde de influir o no en los resultados.

El JU usualmente se usa para medir las preferencias de equidad, por ello es particularmente relevante para las consideraciones de justicia y equidad en otras especies. Hay dos estudios en chimpancés, ambos tomando en cuenta una forma limitada del juego, en el que los que proponen eligen entre dos distribuciones prestablecidas. En ninguno de los dos casos, los que responden rechazaron las distribuciones, sin embargo, la conducta de los que proponen varió a lo largo de los experimentos. En el primero, los que proponen escogieron entre dos bandejas, cada una contenía alimento para él y su compañero (ninguno de los chimpancés podía alcanzar el alimento de su compañero). Después de que el que propone elige una opción, pone a la mano del que responde la bandeja correspondiente; el que responde puede elegir entre quedarse con ella o rechazarla, acercándola al que propone, lo que da lugar a que ambos puedan acceder a esa distribución. En este estudio, los que proponen eligieron consistentemente la distribución más alta y los que responden aceptaron virtualmente cualquier oferta (Jensen, et al. 2007). No obstante, los que responden aceptaron más de la mitad de las distribuciones que no tenían nada (p.ej., ellos no recibieron nada), lo cual indica que probablemente no entendieron la tarea. Un estudio posterior mostró que los humanos que responden rara vez eligen el rechazo en una situación similar (Smith y Silberberg 2010), lo que indica que la desviación de la conducta registrada regularmente en el JU podría atribuirse al diseño experimental.
En un estudio aún más reciente (Proctor, et al. 2013), los sujetos se entrenaron mostrándoles que una ficha representaba una división equitativa de 3/3 piezas a partir de seis pedazos de alimento, mientras que otra ficha representaba una división inequitativa de 5/1 de las piezas de comida a favor del que propone. Los que proponen podían elegir una de las fichas y pasarla al que responde, el cual después podía intercambiarla con el experimentador (con la aceptación de la propuesta) o no (rechazándola). Las elecciones de los que proponen en el JU se compararon con aquellas elecciones en una condición tipo del JD en la que el que responde estaba presente en la interacción pero no tenía juego (los que proponen pasaban la ficha elegida directamente al experimentador). El procedimiento se repitió con niños emparejados con un individuo conocido de su guardería para asegurar que los humanos respondieran de manera similar a los chimpancés en este mismo procedimiento.
A pesar de que los que responden nunca rechazan la oferta, todos los que proponen entre los chimpancés cambiaron de preferir el arreglo más beneficioso, la ficha 5/1 en la condición tipo del JD, a preferir el arreglo equitativo de 3/3 en el JU. Lo que validó el procedimiento fue que los niños respondieron de la misma manera. Estos resultados difieren de los típicos resultados que se obtienen en el JU porque los niños y los chimpancés nunca rechazaron la oferta baja (5/1). Con todo, en el JU típico de los humanos los que responden interactúan con compañeros anónimos, y el único recurso que tienen es el de rechazar la oferta. En este estudio, los que responden interactuaron con individuos de su mismo grupo social y tenían otras opciones además del rechazo, como el castigo o la amenaza del castigo, que es una forma menos costosa de protesta que rechazar un resultado positivo, aunque éste sea relativamente menor (Milinski 2013). De manera similar, en el típico JU los rechazos de los que proponen decrecen cuando se les dan otras opciones [p.ej.,, el que responde le puede escribir una nota al que propone (Xiao y Houser 2005)]. Estos resultados no indican necesariamente que los chimpancés mostraran un sentido de la equidad, ya que si ese fuera el caso, habrían preferido la ficha equitativa tanto en el JD como en el JU. Esto tampoco quiere decir que los que proponen tuvieran motivaciones prosociales, ya que podrían haber cambiado su preferencia para evitar rechazos por parte del que responde. Sin embargo, tanto humanos como chimpancés fueron aparentemente sensibles a cómo sus elecciones influenciaron la decisión de sus compañeros y cambiaron su conducta de acuerdo con ello.

El Juego de la Impunidad
Un juego cercanamente relacionado con el JU es el Juego de la Impunidad (JI) (Fig. 2). El JI es similar al JU, pero si el que responde rechaza la propuesta del que propone, el que responde no se lleva nada, mientras que el que propone recibe su porción de la distribución tal como él mismo lo había dispuesto (Bolton, et al. 1998). Este juego ha tenido muy poca atención dentro de la literatura humana. En parte, esto se desprende de la asunción de que cualquier jugador racional que responde nunca rechaza la oferta, ya que esta acción lo deja con menos que el que propone, tanto en términos relativos como absolutos (en el JU, el rechazo deja al que responde en términos absolutos con menos que el que propone, pero con lo mismo en términos relativos). Aun así, en un estudio que utilizó el mismo procedimiento en ambos juegos, los sujetos humanos rechazaron en el JI alrededor de la mitad de las veces que lo hicieron en el JU para una misma distribución (Yamagishi, et al. 2009).
En primates, tanto en el JU como en el JI, el reto es que los que responden entiendan realmente que su elección está constreñida por las acciones del que propone. La mayoría de los estudios con primates a la fecha han explorado solamente las respuestas a distribuciones inequitativas para aprender cómo es que los individuos responden a la inequidad cuando ellos no pueden alterar los resultados de sus compañeros [revisión en la ref. Price y Brosnan 2012]. En un experimento típico, una pareja de sujetos alterna la realización de una tarea con un experimentador humano (por ejemplo, el intercambio de una ficha). El experimentador recompensa a los sujetos con la misma recompensa (condición Equitativa) o le da a uno la recompensa de mayor preferencia (condición Inequitativa). Para descartar la posibilidad de un efecto contraste (Reynolds 1961; Roma, et al. 2006), en la que los sujetos comparan sus resultados con aquellos presentes en el ambiente sin atender al componente social, las parejas son sometidas a una condición Contraste, en la que a los dos compañeros se les muestra la recompensa preferida pero, después de completar la tarea, reciben la de menor preferencia (Talbot, et al. 2011; Brosnan, et al. 2010). Estas respuestas son referidas como Inequidad y Equidad en vez de Desigualdad e Igualdad, porque independientemente del hecho de que los resultados son idénticos (o no) no podemos saber si son percibidas como idénticas por los sujetos, dadas las diferencias en rango, alimentación, etc.
Los primeros estudios confirmaron que tanto los monos capuchinos (Brosnan y de Waal 2003; van Wolkenten, et al. 2007; Fletcher 2008) como los chimpancés (Brosnan, et al. 2005; Brosnan, et al. 2010) estaban menos dispuestos a completar una interacción (p.ej., regresar la ficha o aceptar la recompensa alimenticia subsecuente) en la condición de Inequidad comparada, tanto con la condición de Equidad, como con la condición Contraste, aunque hubo variación tanto entre los estudios como al interior de los mismos [más adelante se discute en mayor detalle (Brosnan, et al. 2005; Brauer, et al. 2009; Silberberg, et al. 2009)]. Dado que los humanos suelen rechazar con mayor frecuencia en condiciones donde no están de acuerdo con el resultado (p.ej., en el JU), esta conducta similar en otras especies se ha interpretado como una respuesta negativa ante resultados inequitativos. La respuesta es sensible al contexto, con rechazos que ocurren sólo cuando los sujetos han completado la tarea, pero no cuando las recompensas son otorgadas “gratuitamente” (Roma, et al. 2006; Silberberg, et al. 2009; Brauer, et al. 2006; Neiworth, et al. 2009; Dindo y de Waal 2007). A pesar de ello, los sujetos no cambian la tasa de rechazos cuando tienen que trabajar más que un compañero para recibir una recompensa (Fontenot, et al. 2007; Takimoto y Fujita 2011 para una excepción). Esto indica que aunque los sujetos son sensibles a la inequidad en el contexto de la acción conjunta, es la diferencia en el resultado más que el esfuerzo lo que es importante para su toma de decisiones. Esto puede indicar que los sujetos son más sensibles a las inequidades distributivas que a las procesales.
Las variables demográficas también influyen en las respuestas de los individuos, aunque no siempre de manera consistente. Por ejemplo, no todos los estudios muestran un efecto de la dominancia, aunque para aquellos donde sí se observa, los individuos dominantes siempre responden con mayor intensidad que los subordinados, presumiblemente porque están más acostumbrados a recibir la mayor recompensa (Brosnan, et al. 2010; Brauer, et al. 2009). Tampoco hay diferencias sexuales consistentes (Brosnan, et al. 2005; Brosnan, et al. 2010), probablemente por el tamaño relativamente pequeño de las muestras (aunque varias docenas de chimpancés han sido evaluados en tres diferentes instalaciones, lo que constituye una muestra amplia para los estudios con primates cautivos, esta es una muestra muy pequeña para desentrañar las variaciones individuales). El grado al cual las diferencias individuales influye en la conducta —tanto en primates humanos como no humanos— es un asunto que requiere exploración adicional.
Finalmente, no todas las especies de primates rechazan en condiciones de inequidad. El mismo procedimiento para provocar inequidad ha sido implementado en siete especies de primates, y otros procedimientos similares se han utilizado con otras tantas especies (Tabla 1) (Véase www.pnas.org/content/110/supplement-2/10416/T1). La comparación de las respuestas de estas diferentes especies nos permite considerar qué factores sociales y ambientales pueden haber jugado un papel en la modulación de la evolución de la respuesta. Aunque se necesita evaluar a más especies para verificar estas conclusiones, algunos patrones han surgido. Por ejemplo, la respuesta negativa ante la inequidad no es una homología entre los primates. Entre los grandes simios, los bonobos responden de manera similar a los chimpancés y a los humanos, lo cual indica que el último ancestro común de Pan y Homo compartió esta conducta. Solamente cinco bonobos han sido evaluados, lo cual provoca que los análisis estadísticos se compliquen, sin embargo, sus tasas de rechazo se duplicaron de 10 a 20 por ciento en la condición de Inequidad comparada con la de Equidad (Brauer, et al. 2009). Por otra parte, los orangutanes no mostraron ningún tipo de respuesta ante la inequidad (Brauer, et al. 2009; Brosnan, et al. 2011), lo que elimina la posibilidad de una homología entre los grandes simios. Además, esta conducta no es una homología entre los monos del Nuevo Mundo, ya que de los cinco monos del Nuevo Mundo que han sido evaluados sólo los capuchinos respondieron de manera diferente en la condición de inequidad. La conducta de los orangutanes también descarta la posibilidad de que el asunto esté relacionado con un incremento en el tamaño del cerebro o con capacidades cognitivas avanzadas, ya que los orangutanes cuentan con una relación similar del cerebro respecto a la talla y habilidades cognitivas similares a las de los otros grandes simios.
Hasta aquí, la respuesta negativa ante la inequidad se correlaciona positivamente con la posibilidad de una especie para cooperar rutinariamente con individuos no emparentados (revisión en ref. Brosnan 2011) (Fig. 3). Los chimpancés, bonobos y monos capuchinos cooperan con individuos no emparentados, por ejemplo en coaliciones y alianzas, y las tres especies responden a la inequidad en estos experimentos. Los macacos también responden negativamente ante la inequidad (Hopper, et al. 2013; Cronin, et al. 2010) y muestran cooperación en forma de coaliciones y alianzas (Maetripieri 2007). Sin embargo, en los monos ardilla, una especie confamilial (y simpátrica) con los capuchinos pero carente de la extensa cooperación social, la respuesta de los machos tuvo mayor intensidad en la condición de contraste que en las de inequidad y equidad, lo que indica que sus reacciones tienen una mayor influencia por las ofertas previas que por las recompensas recibidas por otro mono (Talbot, et al. 2011). De nuevo, si tomamos en cuenta a los orangutanes, un aspecto en el que difieren de los chimpancés y los bonobos es la relativa carencia de cooperación social. Una dificultad potencial surge entre las especies con cuidado biparental. Estas especies cooperan de manera extensa, con la crianza conjunta de la descendencia, aunque hasta ahora no tenemos indicios de que dichas especies respondan negativamente a la inequidad dentro de este paradigma experimental (Neiworth, et al. 2009). De cualquier forma, la función que se propone acerca de la respuesta ante la inequidad es el juicio sobre el valor de los compañeros que cooperan para determinar la búsqueda de un nuevo compañero. Es muy costoso encontrar un nuevo compañero, con lo cual es muy probable que incluso individuos con un cuidado biparental sean capaces de distinguir las inequidades. Han evolucionado para responder solamente ante inequidades importantes, lo cual no incluye recibir una recompensa menor a la preferida, comparada con la de un compañero.
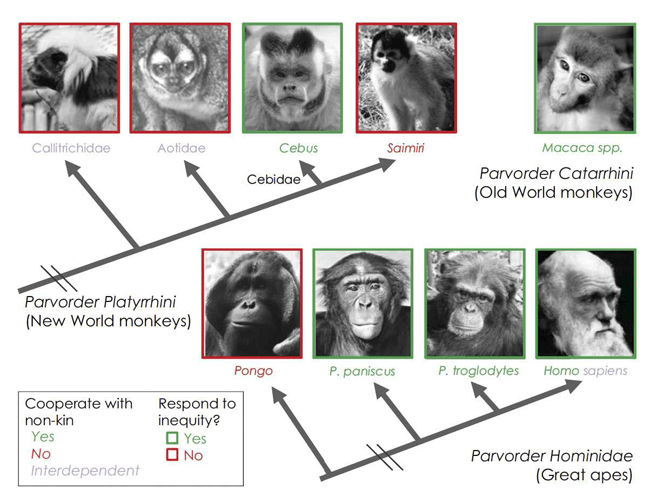
Con relación a esto, los sujetos deben ser más sensibles a la inequidad cuando forman una relación, que es el momento cuando el costo de encontrar a un compañero es menor, que después de que la relación ha sido establecida y hay una inversión conjunta en la descendencia (Brosnan 2011). Un inconveniente de nuestra comprensión de la conducta humana es que la mayoría de los estudios hasta ahora han involucrado individuos no relacionados, la mayoría desconocidos, casi siempre en condiciones de anonimato y frecuentemente sin interacciones repetidas, lo que elimina la posibilidad de formar una reputación. Aunque estas condiciones nos dan información para explorar el grado al que los humanos cooperan sin una relación o sin conocimiento social, esto no nos da información precisa para decirnos algo sobre la conducta cooperativa típica de los humanos, la cual principalmente se presenta al interior de relaciones sociales establecidas con compañeros conocidos. Otros experimentos que analicen las diferencias conductuales en el contexto de diferentes relaciones sociales y con diferentes grados de anonimato son esenciales para la comprensión de cómo funciona la inequidad en el contexto de la relación.
Juegos prosociales
A pesar de los reclamos de los niños de que “eso no es justo” cuando reciben un resultado menor al preferido que otro niño, la respuesta negativa ante la recepción de una menor recompensa que la del compañero es solamente una parte del sentido de la equidad. El sentido de la equidad o la justicia requiere responder no sólo a las inequidades que le afectan a uno mismo, sino también a aquellas que afectan a los demás (Fehr y Schmidt 1999; Hatfield, et al. 1978). Esto ha sido explorado experimentalmente con la utilización de procedimientos reminiscentes del Juego del Dictador. En el típico juego experimental con primates, el sujeto tiene la opción de otorgar recompensas, o no, a un compañero social. Es importante señalar que la conducta del sujeto, cuando está con un compañero social se compara con su conducta cuando está solo, lo que lleva a descartar la posibilidad de que esté eligiendo la opción prosocial, ya que hay una mayor cantidad de recompensas visibles con relación a la opción no prosocial. La diferencia fundamental entre estos juegos y el JD es que la elección de recompensar al compañero no reduce la propia recompensa del sujeto en estos juegos prosociales y, por lo tanto, no hay elementos que desincentiven la conducta prosocial.
Aunque la estructura de las recompensas ha variado a través de los experimentos, lo cual ha resultado en la dificultad de las comparaciones, un número suficiente de estudios de este tipo ha mostrado que hay patrones emergentes. Los chimpancés eligen recompensar a sus compañeros en algunos experimentos (Horner, et al. 2011) pero en otros no (Silk, et al. 2005; Jensen, et al. 2006), y no hay claridad respecto a cuáles características llevan a la conducta prosocial en este simio. Tampoco es claro que estos resultados reflejen las tendencias prosociales de los chimpancés. Los resultados en los estudios de inequidad descritos anteriormente implican que los chimpancés pueden reconocer cuando reciben un mejor resultado que un compañero incluso cuando ellos eligen no modificar la situación. En estos estudios, algunos sujetos rechazaron las recompensas preferidas cuando sus compañeros obtuvieron las menos preferidas y lo hicieron en mayor medida que cuando ambos recibían una recompensa preferida (Brosnan, et al. 2010). Esto no quiere decir que las motivaciones de los sujetos fueran prosociales, ya que pueden haber sentido miedo de la reacción de su compañero al recibir una recompensa menos preferida que él, aunque estos resultados comprueban que los chimpancés son capaces de reconocer cuando reciben una mejor recompensa que sus compañeros.
Entre los monos, los capuchinos eligen preferentemente la opción prosocial (de Waal, et al. 2008; Lakshminarayanan y Santos 2008) y tienen una mayor disposición a recompensar a un individuo emparentado que a individuos no emparentados del mismo grupo o a individuos de otro grupo (de Waal, et al. 2008). Los calitrícidos, uno de los taxa que muestran cuidado biparental, también eligen la opción prosocial que recompensa a un compañero del grupo (Burkart, et al. 2007; Cronin, et al. 2010), aunque al igual que los chimpancés, no lo hacen en todos los experimentos (Cronin, et al. 2009). Por último, los macacos de cola larga también muestran conducta prosocial, si bien en este estudio los dominantes escogen la opción prosocial, mientras que los subordinados no (Massen, et al. 2010). De nuevo, en este estudio se midió la conducta, no sus motivaciones, lo cual no nos dice mucho acerca de por qué los monos se comportan de manera prosocial (p.ej., no nos permiten desentrañar si los sujetos estaban motivados prosocialmente a ayudar a sus compañeros o si tenían una motivación egoísta para evitar repercusiones por no ayudar a sus compañeros). Aunque todavía hay debate respecto de los contextos en los que cada una de estas especies muestra conducta prosocial y las presiones selectivas que la sustenten en algunas circunstancias, todos los primates evaluados hasta ahora eligen activamente un resultado que beneficia a un compañero al igual que a ellos mismos.
APROXIMACIONES ESPECIE-ESPECÍFICOS A LA JUSTICIA Y LA EQUIDAD
Inequidad en el contexto de la cooperación
Si la respuesta a la inequidad es un mecanismo para la identificación de compañeros cooperativos, entonces cuando se requiere cooperación la inequidad puede influir en la conducta. Esto puede explorarse experimentalmente por medio de la introducción de la inequidad en un paradigma experimental de la cooperación. Los monos capuchinos han sido estudiados de manera extensa en el terreno de la cooperación y hay evidencia con relación a que son capaces de comprender las contingencias de una tarea cooperativa y de su sensibilidad a la conducta de sus compañeros (revisión en Brosnan 2010). En el primer experimento, donde se analizó la cooperación en una situación inequitativa, los monos capuchinos podían trabajar juntos para jalar una bandeja y obtener alimento, pero sólo uno de los monos accedía al alimento. Los monos trabajaban juntos solamente si el mono que recibía la comida la compartía con el que no recibió nada. Probablemente con la intención de anticiparse a esto, el mono que recibió el alimento tuvo mayor disposición a compartirlo en esta condición que en una condición controlada, en la cual recibiría el alimento sin la ayuda de su compañero. Lo anterior indica que tienen una sensibilidad a la reacción del compañero y que pueden anticipar la necesidad de ayuda de su compañero (de Waal y Berger 2000).
En un experimento que analizó explícitamente la inequidad y la cooperación, los monos fueron evaluados en un esquema similar en el cual los dos monos siempre recibían alimento, aunque en este caso el alimento difería en el grado de preferencia de los monos. En algunos casos ambos monos recibieron el (mismo) alimento preferido, otras recibieron el (mismo) alimento menos preferido y en otras uno de los monos recibió el alimento preferido, mientras que el otro recibió el menos preferido. Los monos no se encontraban separados ni tenían roles predeterminados por el experimentador, así que en todos los casos los monos debían negociar entre sí desde qué posición jalar la bandeja, que a su vez determinaba quién obtenía cuál de los dos alimentos. El éxito de las parejas al jalar la bandeja no dependió del valor de las recompensas; las parejas jalaban la bandeja con la misma frecuencia para una recompensa con dos alimentos iguales que para una con dos alimentos diferentes. En cambio, los monos mostraron sensibilidad a la conducta de sus compañeros. En algunas parejas los individuos compartieron el acceso al alimento preferido cuando las recompensas eran diferentes, y cada uno recibió esa recompensa aproximadamente la mitad de las veces. Entre estas parejas, el éxito al jalar la bandeja de manera cooperativa fue alto (cerca del 70 por ciento) en las tres condiciones. Por otro lado, en las parejas en las que un miembro dominó la recompensa preferida en la condición de recompensas diferentes, solamente tuvieron éxito al jalar la bandeja alrededor de 30 por ciento de las veces en las tres condiciones diferentes (Brosnan, et al. 2006).
Estos resultados muestran dos aspectos importantes. En primer lugar, los sujetos de las parejas inequitativas aparentemente reaccionaron en contra de su compañero y no en contra de la distribución. Es decir, cuando uno de los miembros dominó la recompensa preferida en la condición de recompensas diferentes, la tasa de ocasiones en las cuales jalaron la bandeja se desplomó en todas las condiciones, incluso en las condiciones donde ambos monos recibían la misma recompensa. Esto apoya la hipótesis de que el reconocimiento de inequidades ayuda a identificar compañeros cooperativos, pues los sujetos rechazaron trabajar con un compañero específico en vez de rechazar un resultado particular. En segundo lugar, entre las parejas donde no hubo dominio de un individuo sobre otro con relación a las recompensas, los sujetos recibieron, por definición, la recompensa menos preferida en la mitad de las ocasiones donde se presentaron recompensas diferentes, y aun así la cooperación se mantuvo. La realidad de la cooperación es que es poco probable que todos los individuos reciban la misma recompensa en cada ensayo, y lo que estos resultados muestran es que los monos capuchinos de alguna manera son capaces de extrapolar a través de múltiples ensayos y así cosechar las ganancias de la cooperación, independientemente de la variación en el resultado.
Inequidad en interacciones con el experimentador
Aunque los datos anteriores se relacionan con la recompensa en primates cuando interactúan con otros primates, ¿cómo reaccionan los primates hacia el experimentador que causó la inequidad? Un estudio reciente examinó con cuál de dos investigadores nuevos los monos preferían interactuar en una tarea en la que el mono le regresa una ficha al experimentador para obtener una recompensa alimenticia. Los sujetos, en primer lugar, tuvieron la experiencia de participar con ambos investigadores, uno de ellos consistentemente otorgó la recompensa ofrecida mientras el otro fracasó en su intento por recompensar a los sujetos con la recompensa ofrecida (o cualquier recompensa) en más del 50 por ciento de los ensayos. Cuando a los monos se les dio la opción de elegir a qué experimentador entregarle la ficha, los sujetos tuvieron la misma disposición a interactuar con ambos experimentadores (Brosnan y de Waal 2009). Sin embargo, puede ser que los monos capuchinos no determinen sus preferencias por uno u otro experimentador basados en la confianza [de hecho, sí muestran preferencias en cuanto a los experimentadores en otras situaciones (Paukner, et al. 2009)], sino que es posible también que la relación entre humanos y primates no humanos en cautiverio pueda influir en los resultados. Independientemente del hecho de que los sujetos en la referencia de Brosnan y de Waal, (2009) nunca interactuaron con los experimentadores que participaron en ese estudio, tuvieron en efecto una amplia experiencia con experimentadores en general, así como con ensayos en donde no se les otorgó ninguna recompensa (p.ej., dada una respuesta incorrecta). Por lo tanto, pueden haber respondido a esta situación con la intención de “corregir” su respuesta, más que reconocer la desconfianza del experimentador. Los perros domésticos, que tienen una relación diferente con los humanos, son capaces de discriminar entre experimentadores en una tarea similar (Horowitz, et al. 2005).
Inequidad en interacciones sociales
Una de las mejores evidencias a favor del sentido de justicia o del sentido de la equidad procede de estudios de observación de las interacciones de primates no humanos en su conducta natural. Estas interacciones, a su vez, ofrecen una oportunidad para mirar más allá de la distribución equitativa. De manera desafortunada, en muchos casos, estos eventos son extremadamente raros, por lo que es difícil realizar señalamientos contundentes acerca de esta conducta. Además, los estudios de observación ofrecen correlaciones que no pueden establecer causalidad sin la implementación de experimentos controlados que pueden manipular una variable a la vez. No obstante, estas situaciones pueden ser la mejor ventana con la que contamos para analizar el sentido de la equidad o de la justicia en primates, ya que son interacciones naturales sin restricciones dadas por las interacciones con los humanos o las contingencias experimentales.
Los chimpancés exhiben un sentido de la “regularidad social” que ha sido relacionado con el sentido humano de la justicia (de Waal 1991). Los chimpancés aparentan tener expectativas sobre la conducta de los demás y se sabe que expresan frustración, en ocasiones de manera agresiva, cuando estas expectativas se violan. Además, dichas respuestas no se limitan al caso de los simios. Tanto los simios como los monos se comprometen con la vigilancia de la conducta en la cual los individuos, usualmente los machos de alto rango, intervienen en las peleas para mantener la estabilidad del grupo. Una de las mejores evidencias que muestran la utilidad de la vigilancia de la conducta procede de estudios realizados con macacos. En estos estudios, la conducta del grupo se comparó en días normales y en días donde el macho dominante se apartó del grupo. Los resultados demuestran la necesidad de los machos por estabilizar las interacciones del grupo (Flack, et al. 2005; Flack, et al. 2006). De manera similar, tanto los machos chimpancés como los macacos, una vez que han establecido su condición dominante intervienen en peleas del lado del perdedor (Watanabe 2001; de Waal 1978). Aunque hay muchas razones para esa conducta, nuevamente indican que estos machos pueden reconocer inequidades sociales en las interacciones de los demás y están dispuestos a actuar en contra de sus propios intereses a corto plazo para corregirlas. Por supuesto, que a largo plazo, mantener la estabilidad del grupo siempre beneficia al macho dominante, con lo que una intervención física puede también enfatizar su poder en el grupo respecto de los demás. No obstante, hay costos a corto plazo por intervenir en una pelea, particularmente a favor de un individuo que es derrotado.
Hace poco tiempo, la inequidad también se relacionó con las peleas lúdicas entre los gorilas. Los gorilas que ganaron la partida en una situación de pelea lúdica se esforzaron por mantener su condición, lo que posiblemente indica que estaban al tanto de la dinámica social y se esforzaron por mantener su ventaja competitiva (van Leeuwen, et al. 2011). Los autores enfatizan el beneficio de estudiar la inequidad en un contexto social natural y consideran cómo las respuestas ante la inequidad varían dependiendo de si el individuo en cuestión tiene la ventaja social. Por razones experimentales prácticas, nuevamente, la mayor parte de los estudios se centra en la inequidad de recompensas alimenticias, aunque la atención ante la inequidad seguramente evolucionó dentro del mundo social y sólo recientemente se adaptó a resultados físicos, como lo puede ser el alimento. Más estudios en el futuro podrán ayudar a determinar cómo la respuesta ante la inequidad varia dependiendo del producto en cuestión.
LA INEQUIDAD Y CONDUCTAS RELACIONADAS
La inequidad interactúa aparentemente con otras conductas, aunque, hasta ahora, ha habido pocos estudios que analicen estas interacciones.
Castigo
El reconocimiento de inequidades es solamente una conducta de muchas otras que apuntalan la cooperación. El castigo, a su vez, puede apoyar la cooperación. Los peces limpiadores, que cooperan con individuos no emparentados durante la limpieza, no responden a la inequidad de recompensas comparado con un compañero social (Raihani, et al. 2012). Sin embargo, los peces se castigan mutuamente si actúan en contra de los intereses de la pareja (Raihani, et al. 2010). Como consecuencia, puede ser que en algunas especies o en situaciones en que encontrar a otro compañero puede dificultarse o ser costoso; el castigo para cambiar la conducta del compañero se utiliza en lugar de encontrar un nuevo compañero (Raihani y McAuliffe 2012).
Inequidad y efectos contraste
Parece probable que la respuesta ante la inequidad comparte mecanismos cognitivos con algunas conductas, como la pérdida de la aversión y efectos contraste, los cuales involucran reacciones ante resultados inesperadamente menores. Lo anterior es muy difícil de discernir experimentalmente dado que la inequidad es esencialmente un efecto contraste con el resultado de otro como referente (de manera similar, la inequidad es una pérdida en referencia al resultado del otro). Sin embargo, se puede comparar la intensidad relativa de las motivaciones utilizando experimentos bien diseñados. Por ejemplo, mientras que algunas especies (chimpancés y monos capuchinos) tuvieron mayor sensibilidad a la inequidad con un compañero que al contraste con resultados ofrecidos previamente, otros (p.ej., los monos ardilla) mostraron lo contrario. El mapeo de dichas preferencias puede ayudar a determinar las influencias subyacentes en la expresión de estas conductas, lo que nos permite intuir la intensidad relativa de los estímulos ambientales sociales y no sociales en la toma de decisiones de una especie.
DIRECCIONES FUTURAS Y CONCLUSIONES
Aunque en las décadas previas hemos aprendido mucho sobre las respuestas a la inequidad en primates no humanos, aun queda mucho más por aprender. En primer lugar, y de manera primordial, los humanos son primates, lo cual hace a los primates no humanos un grupo natural con el cual compararlos, aunque hay otras especies que también responden a la inequidad (Horowitz 2012; Raihani, et al. 2012; Range, et al. 2009; Wascher y Bugnyar 2013). Es necesario más trabajo para identificar qué otras especies responden a la inequidad, si estas respuestas son similares entre especies, y la naturaleza de la respuesta y sus diferencias individuales en estas especies. Esto también dará una prueba más robusta a la hipótesis de que las respuestas a la inequidad se correlacionan con la cooperación.
Otra área importante de investigación es la distinción de situaciones en las que las especies no humanas se dan cuenta de la inequidad en contraste con la respuesta a la inequidad. Todos los estudios antes descritos se centran en las respuestas conductuales ante resultados diferenciales, pero los sujetos pueden percatarse de una situación inequitativa a la cual no responden, por ejemplo, ya que son incapaces de inhibir su deseo por una recompensa, por presión social o por otras razones que todavía no conocemos. Una manera en la cual puede responderse esta pregunta es analizar los cambios afectivos de un individuo a través de diferentes condiciones para identificar aquellas respuestas que se dan en la ausencia de un cambio manifiesto de conducta. Esta aproximación ha resultado exitosa, ya que ha permitido identificar el reconocimiento de inequidades en niños pequeños (LoBue, et al. 2009).
La inequidad se presenta en una variedad de contextos que no se reflejan en la literatura de las especies no humanas, dada la concentración en pruebas experimentales que se preocupan principalmente por los aspectos distributivos. Un elemento crucial de la investigación humana que ha recibido poca atención en la literatura no humana es la inequidad procesal (Skitka 2012). Aun cuando algunas investigaciones preliminares han revelado que no hay cambios en la conducta de los sujetos basada en variaciones procesales (Brosnan, et al. 2010), los primates no humanos tienen expectativas de la conducta de los demás y es razonable asumir que tienen sensibilidad a las diferencias procesales.
Todo el trabajo experimental al día de hoy ha involucrado una aproximación egocéntrica de la equidad y la justicia, aun cuando algunos estudios de observación de la conducta natural indican que los primates, en efecto, intervienen en las interacciones de los demás, en algunas ocasiones de manera imparcial. Aunque es difícil, es posible estudiar la equidad en el laboratorio descartando las reacciones egocéntricas. Los estudios experimentales sobre reputación en chimpancés indican que estos simios son capaces de formarse opiniones de los experimentadores humanos a partir de observarlos en interacciones con otros chimpancés (Subiaul, et al. 2008). Una línea de investigación fértil sería el análisis de los juicios de los primates no humanos sobre la equidad de las interacciones de los demás. Dichas evaluaciones a terceros sobre las distribuciones de los demás serían la primera situación en la cual estudiar experimentalmente la justicia en otras especies.
Finalmente, los estudios sobre justicia y equidad avanzarán de manera significativa cuando el trabajo de laboratorio se combine con lo que sabemos de la conducta de los animales en su ambiente natural. De esta manera, comprenderemos mejor las condiciones sociales y ecológicas bajo las cuales estas preferencias se manifiestan, lo que proporciona conocimiento adicional en términos de su función. Además, la combinación de estos estudios conductuales con lo que sabemos de la cognición puede ayudarnos a comprender estas conductas, tales como la superposición entre la inequidad y el retraso de la gratificación (Price y Brosnan 2012).
Los humanos no son los únicos que responden negativamente al trato diferencial de un compañero. Esta respuesta se comparte con otras especies y aparentemente es básica para lograr una cooperación exitosa. La comprensión de la evolución de esta respuesta nos da luz sobre nuestra propia conducta. Aunque los primates no humanos no muestran un sentido de justicia o equidad de la misma manera que lo hacen los humanos, al acumular conocimiento respecto de la evolución de la justicia y la equidad a partir de estudios comparativos nuestra comprensión de estos juicios mejorará, incluso en lo que tiene que ver con la conducta de los humanos.
AGRADECIMIENTOS
Quiero agradecer a L. Salwiczek por sus útiles comentarios a un borrador previo de este manuscrito. El financiamiento fue otorgado por la Fundación Nacional de Ciencia [National Science Foundation (NSF)], El Galardón al Faculty Early Carrer Development Award SES 0847351 y la beca NSF SES 1123897.
Traducción de Bernardo Yáñez.

Ludus Vitalis, vol. XXI, num. 40, 2013, pp. 249-272.
BIBLIOGRAFÍA
Bolton, G.E., Katok, E. y Zwick, R. (1998), “Dictator game giving: Rules of fairness versus acts of kindnessc International Journal of Game Theory 27:269-299.
Bräuer, J., Call, J. y Tomasello, M. (2006), “Are apes really inequity averse?” Proceedings of the Royal Academy of Science B 273(1605): 3123-3128.
Bräuer, J., Call, J. y Tomasello, M. (2009), “Are apes inequity averse? New data on the tokenexchange paradigma”, American Journal of Primatology 71(2): 175-181.
Brosnan, S.F. y de Waal, F.B.M. (2003), “Monkeys reject unequal pay”, Nature 425(6955): 297-299.
Brosnan, S.F., Schiff, H.C. y de Waal, F.B.M. (2005), “Tolerance for inequity may increase with social closeness in chimpanzees”, Proceedings of the Royal Society B 272(1560): 253-258.
Brosnan, S.F. (2006), “At a crossroads of disciplines”, Social Justice Research 19(2): 218-227.
Brosnan, S.F., Freeman, C. y de Waal, F.B.M. (2006), “Partner’s behavior, not reward distribution, determines success in an unequal cooperative task in capuchin monkeys”, American Journal of Primatology 68(7): 713-724.
Brosnan, S.F. y de Waal, F.B.M. (2009), “Capuchin monkeys tolerate intermittent unreliability in human experimenters”, International Journal of Primatology 30: 663-674.
Brosnan, S.F. (2010), “What Do Capuchin Monkeys Tell Us About Cooperation?” in D.R. Forsyth y C.L. Hoyt(eds.), For the Greater Good of All: Perspectives on Individualism, Society, and Leadership. New York: Palgrave Macmillan Publishers, pp. 11-28.
Brosnan, S.F., et al. (2010), “Competing demands of prosociality and equity in monkeys”, Evolution and Human Behavior 31(4): 279-288.
Brosnan, S.F., Talbot, C., Ahlgren, M., Lambeth, S.P. y Schapiro, S.J. (2010), “Mechanisms underlying the response to inequity in chimpanzees, Pan troglodytes”, Animal Behaviour 79(6): 1229-1237.
Brosnan, S.F., Flemming, T.E., Talbot, C., Mayo, L. y Stoinski, T.S. (2011), “Responses to inequity in orangutans”, Folia Primatologica (Basel) 82: 56-70.
Brosnan, S.F. (2011), “A hypothesis of the co-evolution of inequity and cooperation”, Frontiers in Neuroscience 5: 43.
Brosnan, S.F., Beran, M.J., Parrish, A., Price, S.A. y Wilson, B.J., “Comparative approaches to studying strategy: Towards an evolutionary account of primate decision-making”, Evolutionary Psychology, in press.
Burkart, J.M., Fehr, E., Efferson, C. y van Schaik, C.P. (2007), “Other-regarding preferences in a non-human primate: Common marmosets provision food altruistically”, Proceedings of the National Academy of Sciences USA 104(50): 19762-19766.
Camerer, C. (2003), Behavioral Game Theory: Experiments in Strategic Interaction. Princeton: Russell Sage Foundation; Princeton Univ Press.
Cronin, K.A., Schroeder, K.K.E. y Snowdon, C.T. (2010), “Prosocial behaviour emerges independent of reciprocity in cottontop tamarins”, Proceedings of the Royal Society B 277(1701): 3845-3851.
de Waal, F.B.M. (1978), “Exploitative and familiarity-dependent support strategies in a colony of semi-free living chimpanzees”, Behaviour 66: 268-312.
de Waal, F.B.M. (1991), “The chimpanzee’s sense of social regularity and its relation to the human sense of justice”, American Behavioral Scientist 34(3): 335-349.
de Waal, F.B.M. y Berger, M.L. (2000), “Payment for labour in monkeys”, Nature 404(6778): 563.
de Waal, F.B.M., Leimgruber. K. y Greenberg, A.R. (2008), “Giving is self-rewarding for monkeys”, Proceedings of the National Academy of Sciences USA 105(36): 13685-13689.
Dindo, M. y de Waal, F.B.M. (2007), “Partner effects on food consumption in brown capuchin monkeys”, American Journal Primatology 69(4): 448-456.
Eisenberg Nancy y Mussen P. Henry (1989), The Roots of Prosocial Behavior in Children. Cambridge Univ Press, Cambridge, UK; New York.
Fehr, E. y Schmidt, K.M. (1999), “A theory of fairness, competition, and cooperation”, Quarterly Journal of Economics 114: 817-868.
Flack, J.C., de Waal, F.B.M. y Krakauer, D.C. (2005), “Social structure, robustness, and policing cost in a cognitively sophisticated species”, American Naturalist 165(5): E126-E139.
Flack, J.C., Girvan, M., de Waal, F.B.M. y Krakauer, D.C. (2006), “Policing stabilizes construction of social niches in primates”, Nature 439(7075): 426-429
Fletcher, G.E. (2008), “Attending to the outcome of others: Disadvantageous inequity aversion in male capuchin monkeys (Cebus apella)”, American Journal of Primatology 70(9): 901-905.
Fontenot, M.B., Watson, S.L., Roberts, K.A. y Miller, R.W. (2007), “Effects of food preferences on token exchange and behavioural responses to inequality in tufted capuchin monkeys, Cebus apella”, Animal Behaviour 74(3): 487-496.
Frank, Robert H. (1988), Passions Within Reason: The Strategic Role of the Emotions. New York, W. W. Norton.
Guth, W., Schmittberger, R. y Schwartze, B. (1982), “An experimental analysis of ultimátum bargaining”, Journal of Economic Behavior and Organization 3: 367388.
Hatfield, E., Walster, G.W. y Berscheid, E. (1978), Equity: Theory and Research. Boston: Allyn and Bacon.
Henrich Joseph Patrick et al. (2004) Foundations of Human Sociality: Economic Experiments and Ethnographic Evidence from Fifteen Small-ScaleSocieties. Oxford Univ Press.
Hopper, L.M., Lambeth, S.P., Schapiro, S.J., Bernacky, B.J. y Brosnan, S.F. (2013), “The ontogeny of social comparisons in rhesus macaques (Macaca mulatta)”, Journal of Primatology 2: 109.
Horner, V., Carter, J.D., Suchak, M. y de Waal, F.B.M. (2011), “Spontaneous prosocial choice by chimpanzees”, Proceedings of the National Academy of Sciences USA 108(33): 13847-13851.
Horowitz, A. (2012), “Fair is fine, but more is better: Limits to inequity aversion in the domestic dog’, Social Justice Research 25(2): 195-212.
Jensen, K., Hare, B., Call, J. y Tomasello, M. (2006), “What’s in it for me? Self-regard precludes altruism and spite in chimpanzees”, Proceedings of the Royal Society B 273(1589): 1013-1021.
Jensen, K., Call, J. y Tomasello, M. (2007), “Chimpanzees are rational maximizers in an ultimatum game”, Science 318(5847): 107-109.
Lakshminarayanan, V.R. y Santos, L.R. (2008), “Capuchin monkeys are sensitive to others’ welfare”, Current Biology 18(21): R999-R1000.
LoBue, V., Nishida, T., Chiong, C., DeLoache, J.S. y Haidt, J. (2009), “When getting something good is bad: Even three-year-olds react to inequality”, Social Development 20(1): 154-170.
Maestripieri, D. (2007), Macachiavellian Intelligence. Chicago: Univ. of Chicago Press.
Massen, J.J.M., van den Berg, L.M., Spruijt, B.M. y Sterck, E.H.M. (2010), “Generous leaders and selfish underdogs: Pro-sociality in despotic machaques”, PLoS ONE 5(3): e9734.
Massen, J.J.M., Van Den Berg, L.M., Spruijt, B.M. y Sterck, E.H.M. (2012), “Inequity aversion in relation to effort and relationship quality in long-tailed Macaques (Macaca fascicularis)”, American Journal of Primatology 74(2): 145-156.
Milinski, M. (2013), “Chimps play fair in the ultimatum game”, Proceedings of the National Academy of Sciences USA 110(6): 1978-1979.
Neiworth, J.J., Johnson, E.T., Whillock, K., Greenberg, J. y Brown, V. (2009), “Is a sense of inequity an ancestral primate trait? Testing social inequity in cotton top tamarins (Saguinus oedipus)”, Journal of Comparative Psychology 123(1): 10-17.
Paukner, A., Suomi, S.J., Visalberghi, E. y Ferrari, P.F. (2009), “Capuchin monkeys display affiliation toward humans who imitate them”, Science 325(5942): 880-883.
Price, S.A. y Brosnan, S.F. (2012), “To each according to his need? Variability in the responses to inequity in non-human primates”, Social Justice Research 25(2): 140-169.
Proctor, D., Williamson, R.A., de Waal, F.B.M. y Brosnan, S.F. (2013), “Chimpanzees play the ultimatum game”, Proceedings of the National Academy of Sciences USA 110(6): 2070-2075.
Reynolds, G.S. (1961), “Behavioral contrast”, Journal of the Experimental Analysis of Behavior 4: 441-466.
Raihani, N.J., Grutter, A.S. y Bshary, R. (2010), “Punishers benefit from third- party punishment in fish”, Science 327(5962): 171.
Raihani, N.J., McAuliffe, K., Brosnan, S.F. y Bshary, R. (2012), “Are cleaner fish (Labroides dimidiatus) inequity averse?” Animal Behaviour 84(3): 665-674.
Raihani, N.J. y McAuliffe, K. (2012), “Does inequity aversion motivate punishment? Cleaner fish as a model system”, Social Justice Research 25(2): 213-231.
Range, F., Horn, L., Viranyi, Z. y Huber, L. (2009), “The absence of reward induces inequity aversion in dogs”, Proceedings of the National Academy of Sciences USA 106(1): 340-345
Roma, P.G., Silberberg, A., Ruggiero, A.M. y Suomi, S.J. (2006) “Capuchin monkeys, inequity aversion, and the frustration effect”, Journal of Comparative Psychology 120(1): 67-73.
Silberberg, A., Crescimbene, L., Addessi, E., Anderson, J.R. y Visalberghi, E. (2009), “Does inequity aversion depend on a frustration effect? A test with capuchin monkeys (Cebus apella)”, Animal Cognition 12(3): 505-509.
Silk, J.B., et al. (2005), “Chimpanzees are indifferent to the welfare of unrelated group members”, Nature 437(7063): 1357-1359.
Skitka, L.J. (2012), “Cross-disciplinary conversations: A social psychological perspective on justice research with non-human animals”, Social Justice Research 25(3): 327-335.
Smith, P. y Silberberg, A. (2010), “Rational maximizing by humans (Homo sapiens) in an ultimatum game”, Animal Cognition 13(4): 671-677.
Smith, V.L. (1987), “Experimental methods in economics”, in J. Eatwell, M. Milgate y P. Newman (eds.) The New Palgrave: A Dictionary of Economics. London: Macmillan, pp. 241-248.
Subiaul, F., Vonk, J. Okamoto-Barth, S. y Barth, J. (2008), “Do chimpanzees learn reputation by observation? Evidence from direct and indirect experience with generous and selfish strangers”, Animal Cognition 11(4): 611-623.
Takimoto, A., Kuroshima, H. y Fujita, K. (2010), “Capuchin monkeys (Cebus apella) are sensitive to others’ reward: An experimental analysis of food-choice for conspecifics”, Animal Cognition 13(2): 249-261.
Takimoto, A. y Fujita, K. (2011), “I acknowledge your help: Capuchin monkeys’ sensitivity to others’ labor”, Animal Cognition 14(5): 715-725.
Talbot, C.F., Freeman, H.D., Williams, L.E. y Brosnan, S.F. (2011), “Squirrel monkeys’ response to inequitable outcomes indicates a behavioural convergence within the primates”, Biology Letters 7(5): 680-682,
10.1098/rsbl.2011.0211.
van Leeuwen, E., Zimmermann, E. y Davila Ross, M. (2011), “Responding to inequities: Gorillas try to maintain their competitive advantage during play rights”, Biology Letters 7(1): 39-42.
van Wolkenten, M., Brosnan, S.F. y de Waal, F.B.M. (2007), “Inequity responses of monkeys modified by effort”, Proceedings of the National Academy of Sciences USA 104(47): 18854-18859.
Wascher, C.A.F. y Bugnyar, T. (2013), “Behavioral responses to inequity in reward distribution and working effort in crows and ravens”, PLoS ONE 8(2): e56885.
Watanabe, K. (2001), “A review of 50 years of research on the Japanese monkeys of Koshima: Status and dominante,” in T. Matsuzawa (ed.), Primate Origins of Human Cognition and Behavior. Tokyo: Springer, pp. 405-417.
Wilson, B.J. (2012) “Contra Private Fairness”, American Journal of Economics and Sociology 71(2): 407-435.
Xiao, E. y Houser, D. (2005), “Emotion expression in human punishment behavior”, Proceedings of the National Academy of Sciences USA 102(20): 7398-7401.
Yamagishi, T., et al. (2009), “The private rejection of unfair offers and emotional commitment”, Proceedings of the National Academy of Sciences USA 106(28): 11520-11523.