La Similitud en la Forma y la Función del Hipocampo en Roedores, Monos y Humanos
Robert E. Clark(a,b), Larry R. Squire(a,b,c,f)
ABSTRACT. We begin by describing an historical scientific debate in which the fundamental idea that species are related by evolutionary descent was challenged. The challenge was based on supposed neuroanatomical differences between humans and other primates with respect to a structure known then as the hippocampus minor. The debate took place in the early 1860s, just after the publication of Darwin’s famous book. We then recount the difficult road that was traveled to develop an animal model of human memory impairment, a matter that also turned on questions about similarities and differences between humans and other primates. We then describe how the insight that there are multiple memory systems helped to secure the animal model and how the animal model was ultimately used to identify the neuroanatomy of longterm declarative memory (sometimes termed explicit memory). Finally, we describe a challenge to theanimal model and to cross-species comparisons by considering the case of the concurrent discrimination task, drawing on findings from humans and monkeys. We suggest that analysis of such cases, based on the understanding that there are multiple memory systems with different properties, has served to emphasize the similarities in memory function across mammalian species.
Describiremos aquí el exitoso desarrollo durante las últimas décadas de un modelo de la memoria humana y del deterioro de la memoria humana llevado a cabo en monos y roedores. Este trabajo, que se enfocó en el hipocampo y las regiones cerebrales anatómicamente relacionadas, ofrece una ilustración sólida de una neuroanatomía y funciones similares a través de las especies. La expectativa de que el estudio experimental en animales debe ayudar a comprender la organización de la memoria humana es ciertamente razonable cuando se le considera a la luz de la evolución y la teoría de la evolución. En efecto, la idea central detrás del estudio de los mamíferos no humanos es la noción de que lo que se puede aprender sobre el comportamiento y el cerebro de los mamíferos puede mejorar el entendimiento del cerebro humano y de la condición humana. No fue tan fácil alcanzar esta perspectiva. Comenzaremos por describir un debate científico histórico en el que esta idea fundamental (la continuidad entre no humanos y humanos) fue cuestionada sobre la base de supuestas diferencias neuroanatómicas entre los humanos y otros primates. El debate tuvo lugar a principios de 1860, justo después de la publicación, en 1859, de El origen de las especies de Darwin (1859). A continuación expondremos el difícil camino que se recorrió para desarrollar un modelo animal del deterioro de la memoria humana.
EL HIPOCAMPO MENOR Y LA ANATOMÍA DEL CEREBRO
En 1859, sir Richard Owen (1804-1892), el renombrado anatomista comparativo y paleontólogo británico, superintendente del Departamento de Historia Natural del Museo Británico, miembro de la Real Sociedad y autor de más de seiscientas publicaciones científicas, publicó un provocativo y controvertido trabajo (Owen 1859). Owen argumentó que los humanos debían ser reclasificados como distintos y separados de otros primates. Hasta ese momento, la clasificación de Linneo había ubicado a los humanos, los simios y los monos en un solo orden, el de los primates. La nueva clasificación de Owen no sólo creó un nuevo orden para los seres humanos (Bimana “los de dos manos”), sino una nueva subclase de humanos (Archencephala “cerebro dominante”), que ubica a los humanos muy aparte de otros primates. En la página final de su ensayo, Owen provee el siguiente resumen: “El hombre es la única especie de su género, el único representante de su orden y subclase. Así que yo confío en la refutación que se ha presentado de la transformación del mono en hombre, que aparece en un viejo y favorito autor, que ha sido bien recibido por algunos en su día” (Owen 1859).
Se ha argumentado (Gross 1993a) que esta reclasificación pudo haber sido debido, al menos en parte, al conocimiento de Owen de la próxima publicación de Charles Darwin (1859) de El origen de las especies. Era bien sabido, incluso antes de su publicación, que el libro de Darwin enfatizaría una clara relación fisiológica y anatómica entre las especies y sostendría que todas las especies descienden de un tronco común (además de proponer la teoría de la “selección natural” como el mecanismo para la conducción de la evolución).
Owen fundó su nueva clasificación en tres diferencias neuroanatómicas ostensibles en la estructura cerebral entre los humanos y otros animales, que incluye a los primates no humanos. Owen sugirió que estos tres rasgos anatómicos eran fundamentales, porque estas estructuras no eran simplemente diferentes en los humanos y otros primates, sino que se encontraban únicamente en el cerebro humano (Owen 1859). De acuerdo con Owen, la primera diferencia fundamental entre los humanos y los animales era que sólo en los humanos había un lóbulo posterior. Más precisamente, sólo el cerebro humano se extendió sobre y más allá del cerebelo. La segunda diferencia era que sólo los humanos tienen un cuerno o asta posterior en sus ventrículos laterales (una porción del ventrículo lateral que se extiende posterior y lateralmente). La tercera y más importante diferencia era que sólo los humanos tienen un hipocampo menor.
En la base de los ventrículos laterales hay dos estructuras prominentes: una gran estructura referida en ese momento como el hipocampo mayor (ahora conocido simplemente como el hipocampo) y una estructura más pequeña conocida entonces como el hipocampo menor. El hipocampo menor es una cordillera en el piso del cuerno posterior del ventrículo lateral. El término original para el hipocampo menor, y el término de uso corriente, es el calcar avis. El calcar avis es un término latino que significa “espolón de gallo”, debido a su parecido con las espuelas que se encuentran en las patas de un gallo.
La reclasificación de Owen se encontró con una fuerte oposición, liderada principalmente por Thomas Henry Huxley (1825-1895). Huxley fue un biólogo británico, paleontólogo y anatomista. Durante su vida se desempeñó como presidente de la Real Sociedad, presidente de la Sociedad Geológica y presidente de la Asociación Británica para el Avance de la Ciencia, y era uno de los más elocuentes partidarios científicos de Darwin. En cierto sentido, el debate entre Owen y Huxley fue poco emocionante, ya que Huxley pudo fácilmente demostrar que los tres criterios de Owen, que incluía el del hipocampo menor, no sólo eran identificables en todas las especies de primates disponibles para el estudio, sino que el tamaño relativo del hipocampo menor era más grande en algunas especies de primates que en los humanos (Huxley 1861a, 1861b, 1863). Durante el curso de sus investigaciones, Huxley descubrió también que la fisura del hipocampo en primates en realidad consistía en dos fisuras separadas. La fisura más anterior mantuvo el nombre de fisura del hipocampo, mientras que la fisura más posterior fue renombrada por Huxley (1861b) como surco calcarino porque el calcar avis, o hipocampo menor, es, de hecho, formado por una importante penetración hacia el interior de la surco calcarino. El debate sobre el hipocampo menor, se puede decir, concluyó cuando sir Charles Lyell, un geólogo destacado de la Gran Bretaña y uno de los científicos más eminentes, examinó en detalle el debate del hipocampo menor (Lyell 1863). Lyell con autoridad llegó a la conclusión, sin reservas, de que el debate sobre el hipocampo menor se resolvía a favor de Huxley. La conclusión de Lyell era tanto más convincente porque él no había adoptado completamente la teoría de la evolución de Darwin.
A pesar de su corta vida, el debate sobre el hipocampo menor fue importante. Algunos historiadores (Desmond 1984, Gross 1993b) han sugerido que Huxley intentó mostrar no sólo que la teoría de Owen era incorrecta, sino que debió haber oscurecido los hechos de manera intencional. Huxley lo admitió en una carta que envió a sir Charles Lyell el 17 de agosto de 1862 (la carta se reproduce en Huxley 1900). En esta carta, Huxley hace referencia a un verso de un poema muy conocido señalado por el destacado paleontólogo sir Philip Egerton. Huxley escribió: “No creo que usted encuentre espacio para quejarse de cualquier falta de claridad en mi definición de la posición de Owen con relación a la cuestión del hipocampo. Me refiero a ofrecer la historia completa del asunto en una nota que debe estar fuera de toda duda, la nota parafrasea la línea de sir Egerton ’A lo que Huxley responde que Owen miente’ “. El esfuerzo de Huxley dañó la reputación científica de tal vez el más prominente antievolucionista de la época y ayudó a preparar el terreno para la amplia aceptación científica de la teoría de la evolución por los científicos durante las próximas décadas (se ofrecen excelentes revisiones del debate sobre el hipocampo menor en Gross 1993a, 1993b).
Al final, el debate sobre el hipocampo menor proporcionó otro ejemplo de las similitudes en la organización neuroanatómica entre todas las especies de primates, en las que se incluye a los humanos. Más tarde se probarían estas similitudes para la función del hipocampo y para la organización de la memoria a largo plazo en las especies que van desde los roedores a los primates. Sin embargo, como se describe en las siguientes secciones, la continuidad del funcionamiento de la memoria a lo largo de las especies se mantuvo seriamente en duda durante casi dos décadas.
EL HIPOCAMPO Y LA ERA MODERNA EN LA INVESTIGACIÓN DE LA MEMORIA
La era moderna en la investigación de la memoria, así como las primeras ideas sobre el hipocampo y la organización de la memoria, se inició con la descripción del paciente H. M. por William Beecher Scoville y Brenda Milner (1957). El paciente H. M. tenía un historial de convulsiones menores y mayores que no respondían a la medicación antiepiléptica. Finalmente, con el consentimiento de la familia, el cirujano Scoville trató de aliviar las convulsiones con una intervención quirúrgica experimental que implicaba cortar la cara medial del lóbulo temporal bilateralmente. Posteriormente, la resonancia magnética mostró que la lesión fue bilateralmente simétrica e incluía la cara medial de la corteza polo temporal, prácticamente toda la corteza entorrinal, el complejo amigdaloide y la mitad anterior de la cara intraventricular de la formación del hipocampo [es decir, el giro dentado (GD), el hipocampo y el complejo subicular]. Del mismo modo, la corteza perirrinal fue sustancialmente dañada, con pocas limitaciones en su aspecto ventrocaudal (Corkin, et al. 1997). La cirugía tuvo éxito en la reducción de la frecuencia y la gravedad de las convulsiones, pero dejó a H. M. con una profunda amnesia.
La posterior evaluación sistemática de H. M. y otros pacientes con daño similar establecieron tres principios fundamentales (Squire 2009). En primer lugar, la memoria es una habilidad cerebral específica que está separada de otras funciones cognitivas, tales como la percepción, la inteligencia, la personalidad y la motivación. En segundo lugar, la memoria a corto plazo y la memoria a largo plazo son funciones distintas, porque H. M. tenía un deterioro severo de la memoria a largo plazo. Sin embargo, podía mantener y utilizar la información por un corto periodo de tiempo en la memoria inmediata (y en la memoria de trabajo) siempre que el material pudiera ser ensayado con eficacia. La información se perdía con la distracción. En tercer lugar, las estructuras mediales del lóbulo temporal no son el depósito final de la memoria a largo plazo, puesto que la memoria de H. M. para eventos remotos permaneció en gran parte intacta.
En el momento que H. M. fue descrito por primera vez, la anatomía del lóbulo temporal medial era poco conocida y no se sabía qué daño concreto (dentro de la gran región incluida en su cirugía) era la responsable del deterioro de la memoria. En consecuencia, se iniciaron esfuerzos para abordar esta cuestión en animales de experimentación. De hecho, estos esfuerzos comenzaron casi inmediatamente cuando el propio Scoville fue a Montreal y realizó en monos la misma cirugía que había realizado con H. M. (Correll y Scoville 1965). Sin embargo, éstos y otros monos con lesiones en el lóbulo temporal medial fueron capaces de aprender tareas que parecían similares a las tareas que H. M. no podía aprender.
La dificultad consistía en que todavía no se reconocía que los humanos y los animales de experimentación pueden acercarse claramente a tareas similares haciendo uso de diferentes estrategias (múltiples sistemas de memoria). Por ejemplo, los monos aprenden tareas de discriminación visual de manera gradual a lo largo de muchos ensayos, del modo que ahora se conoce como el ‘aprendizaje de hábitos’. En el mono, este tipo de aprendizaje depende de los ganglios basales en lugar del lóbulo temporal medial (Mishkin, Malamut y Bachevalier 1984; Teng, et al. 2000). Por el contrario, los humanos aprenden la misma tarea cuando se memoriza directamente el material de estímulo. En la década siguiente a la descripción inicial de H. M., un extenso trabajo en ratas con lesiones del hipocampo también falló en reproducir el deterioro de H. M.. Por ejemplo, las ratas con lesiones del hipocampo, así como los monos, ejecutaron de manera normal problemas de discriminación visual simple (Kimble 1963). Las ratas lesionadas tampoco tuvieron deficiencias para aprender a presionar una barra para obtener comida y para aprender diversas tareas en la que evitaban choques (Douglas 1967). En resumen, del trabajo llevado a cabo durante la década de 1960 estaba claro que los deterioros conductuales observados en ratas con lesiones en el hipocampo no ofrecían una descripción adecuada del deterioro en la memoria visto en humanos con lesiones del hipocampo. En consecuencia, los investigadores que estudiaban las ratas no buscaban relacionar su trabajo con los estudios de pacientes con trastornos de memoria y eran más propensos a interpretar sus resultados dentro del marco de la inhibición de respuesta como fue esbozado por primera vez en Pavlov (1927). De hecho, en una revisión de la literatura sobre el hipocampo y el comportamiento publicada una década después de la descripción de H. M., Robert Douglas (1967) escribió:
Las lesiones del hipocampo, obviamente, no conducen a un deterioro en el aprendizaje en general, incluso cuando el aprendizaje implica la retención durante largos periodos de tiempo. Así, los datos humanos y animales podrían parecer contradictorios. Esta contradicción podría ser ’resuelta’, si se postula que el hipocampo tiene una función básica diferente en el hombre y la bestia. Sin embargo, tal solución a este problema es, en general, inaceptable para los fisiopsicólogos. Otra posible solución de esta paradoja es que la reciente pérdida de la memoria en el hombre es un efecto secundario de un tipo diferente de trastorno primario. El autor ha elegido este último supuesto, y sugiere que la reciente pérdida de la memoria en el hombre es un fenómeno genuino, pero que es un subproducto de la interferencia durante el almacenamiento y no debido a una falta per se de la capacidad de almacenar.
Estos comentarios dejaron en claro que una gran parte de la literatura en animales estaba sustancialmente fuera de registro con el trabajo humano y que los experimentadores estaban empezando a dudar de la idea básica de que el daño del lóbulo temporal medial producía un deterioro en la memoria. Además, si uno realmente supone que el daño del lóbulo temporal medial en los humanos causa deterioros en la memoria, entonces, para entender los datos de los animales, se debe considerar la posibilidad, aun cuando sea insatisfactoria, de que la memoria esté organizada de forma diferente en los humanos y en otros animales.
LOS MÚLTIPLES SISTEMAS DE MEMORIA
Durante las décadas de 1960 y 1970 todavía no se entendía que las diferentes tareas de aprendizaje y memoria podrían estar fundadas en diferentes sistemas cerebrales. Muchas de las tareas presentadas a los animales con lesiones del hipocampo eran, de hecho, tareas basadas en habilidades que los pacientes amnésicos habrían podido adquirir (Milner 1962) o bien eran tareas que los animales podían aprender como una habilidad, aun si los humanos tendían a aprender la tarea memorizando el material. El establecimiento de un modelo animal del deterioro de la memoria humana requeriría el desarrollo de tareas para los animales que evaluaran el mismo tipo de memoria que está deteriorada en los humanos después del daño en el lóbulo temporal medial.
El tipo de memoria que está deteriorada es hoy día conocida típicamente como la memoria declarativa (Cohen y Squire 1980). La memoria declarativa se expresa flexiblemente y proporciona acceso consciente a hechos y eventos. Esta memoria se deteriora cuando las estructuras del lóbulo temporal medial se dañan. Por el contrario, el término de la memoria no declarativa hace referencia a diversas destrezas y habilidades adquiridas que no son accesibles al conocimiento consciente, aunque se expresan a través de la ejecución y dependen de diferentes sistemas del cerebro (Squire 2004). La memoria no declarativa es independiente del lóbulo temporal medial.
Un avance clave en el establecimiento de un modelo de la amnesia del lóbulo temporal medial en el ser humano fue el desarrollo para monos de pruebas de memoria en un solo ensayo (one-trial memory tests). Gaffan (1974) sugirió que muchas pruebas de memoria en animales con lesión del hipocampo podían no ser similares a las pruebas que revelan deterioro de la memoria en pacientes humanos. Así pues, si uno intenta relacionar el trabajo en animales con el trabajo en humanos, no es adecuado utilizar cualquier tarea práctica en la que un animal tenga que utilizar la memoria. Más bien, es necesario hacer uso de “pruebas análogas diseñadas específicamente para animales de aquellas pruebas que revelan deterioro en humanos amnésicos” (Gaffan 1974). Una de las pruebas que Gaffan utilizó fue una prueba en un solo ensayo de reconocimiento de memoria visual, que había sido perfeccionada para su uso en monos por Benjamin Weinstein (1941). En esta prueba (conocida inicialmente como el “procedimiento de reacción retardada de igualación de la muestra (matching-from-sample)”), los monos fueron presentados con un solo objeto que sustituían por comida como premio (la fase de muestra). La memoria para el objeto de la muestra fue entonces puesta a prueba después de un retraso, presentando el objeto de la muestra junto con un nuevo objeto (fase de elección). El mono fue entrenado para seleccionar (igualar) el objeto que se había presentado previamente durante la fase de muestra. Se usaron objetos únicos en cada ensayo, por lo que el juicio de familiaridad fue suficiente para identificar el objeto correcto. La demanda de memoria se puede aumentar con el aumento del retardo entre las fases de muestra y la de elección. Esta tarea se conoce como la “igualación retrasada con la muestra” (delayed matching to sample) (IRM). Los monos con lesiones del fórnix, pensados para imitar los efectos de daño del hipocampo, tuvieron deterioros en la ejecución de un modo que dependía de retraso. La ejecución estaba intacta en un retraso de 10 segundos, pero deteriorada en retrasos de 70 y 130 segundos (Gaffan 1974).
Se desarrolló también una versión alternativa de la tarea IRM, conocida como la tarea de falta de igualación retardada con la muestra (FIRM) (Mishkin y Delacour 1975). En este caso, los monos fueron entrenados para seleccionar el nuevo objeto en la fase de selección (i.e., la falta de igualación con el objeto de la muestra). En un experimento crucial (Mishkin 1978), los monos recibieron lesiones diseñadas para imitar los daños sufridos por H.M. El grupo clave recibió daños en el hipocampo además de lesiones en la amígdala, junto con la corteza que subyace a estas dos estructuras. Otros monos recibieron lesiones más pequeñas en el hipocampo (y la corteza subyacente) o lesiones en la amígdala (y la corteza subyacente). Hay que tener en cuenta que las áreas corticales subyacentes, dañadas por la cirugía, no eran el objetivo específico, pero fueron dañadas durante la intervención quirúrgica al hipocampo y la amígdala. Después de la operación, los animales recuperaron la regla de falta de igualación, y la demora entre las fases de muestra y de elección fue incrementada de manera progresiva de 10 a 30, 60 y 120 segundos. El resultado fue que las lesiones del hipocampo o la amígdala (combinado con las áreas corticales subyacentes que fueron dañadas en este estudio) produjeron sólo un leve deterioro, pero la lesión combinada (hipocampo más amígdala y la corteza subyacente) produjo un marcado déficit dependiente del retraso, que fue especialmente severo en demoras más prolongadas.
La demostración de los deterioros dependientes del retraso en la ejecución fue fundamental para la identificación de esta alteración como un deterioro de la memoria. Cuando la ejecución se divide en retrasos cortos y se deteriora selectivamente en retrasos más largos (cuando la demanda de la memoria es mayor), se puede descartar una variedad de explicaciones alternativas para el deterioro (por ejemplo, la capacidad de identificar objetos; los cambios motivacionales; así como efectos secundarios de la lesión, tales como hiperactividad, aumento de la distracción, trastornos motores y otros efectos no específicos). Este estudio, y algunos posteriores, dependientes sobre todo a la tarea FIRM (Zola-Morgan, Squire y Mishkin 1982), documentaron el establecimiento exitoso de un modelo animal de la amnesia del lóbulo temporal medial en el humano.
La tarea FIRM con estímulos de ensayo único fue adaptada posteriormente para su uso con ratas (Mumby, Pinel y Wood 1990). Esta nueva tarea utiliza objetos de ensayo único que podían ser sustituidos para obtener comida como recompensa, al igual que en la versión desarrollada para los monos (Mumby, Pinel y Wood 1990). Además, la prueba fue diseñada de modo que el retraso entre las fases de muestra y de elección podría ser tan corto como 4 segundos. El trabajo con esta tarea demostró que las lesiones del hipocampo, o las lesiones de las regiones corticales adyacentes al hipocampo, producían un deterioro dependiente de retraso en la memoria, similar a lo que se había reportado en el mono (Mumby y Pinel 1994).
Por lo tanto, esta nueva tarea extendió con éxito el modelo animal del deterioro de la memoria del lóbulo temporal medial para la rata. El trabajo adicional con esta tarea demostró que el deterioro en ratas con lesiones del hipocampo era inequívocamente un deterioro de la memoria dependiente de retraso (Clark, West, Zola y Squire 2001) y que la dependencia de retraso no era una simple consecuencia de la gran cantidad de entrenamiento dado en el retraso corto durante el aprendizaje de la regla de falta de igualación (Ringo 1993).
Desde el establecimiento exitoso de la tarea FIRM, otra tarea de la memoria ha sido ampliamente utilizada. Esta tarea permite el estudio del aprendizaje en un solo ensayo en humanos y animales de experimentación de una manera directa y sencilla (Clark y Squire 2010). La tarea visual de comparación pareada (VCP) fue desarrollada inicialmente para el estudio del desarrollo de la visión en los infantes humanos (Fagan 1970). En esta tarea, dos estímulos idénticos son presentados lado a lado durante un periodo de tiempo (por ejemplo, de 1 a 2 minutos). Inmediatamente después, se presenta al infante el estímulo visto recientemente junto con un nuevo estímulo. El descubrimiento es que los infantes prefieren mirar el estímulo nuevo, presumiblemente porque recuerdan el estímulo ya presentado que resulta ahora menos interesante. Este trabajo estableció que la tarea VCP es una prueba fiable de la memoria de reconocimiento visual. La tarea aprovecha una preferencia espontánea por la novedad (que se comparte a través de las especies de mamíferos) y tiene la ventaja de no requerir ninguna instrucción verbal o aprendizaje de reglas. Este rasgo hace de la tarea una excelente herramienta para el estudio de infantes humanos preverbales y animales de experimentación.
La prueba VCP fue posteriormente adaptada para el mono (Bachevalier, Brickson y Hagger 1993), la rata (Ennaceur y Delacour 1988) y el ratón (Tang, et al. 1999). Cuando se utiliza con los primates, esta tarea se denomina generalmente como la tarea VCP, debido a que es una prueba de la memoria de reconocimiento visual. En la rata y el ratón, el animal explora físicamente los objetos de manera que la información visual, olfativa y táctil puede ser utilizada para guiar la ejecución en la fase de prueba. Por eso, en el roedor, esta tarea es típicamente llamada junto con otros descriptores como “reconocimiento del objeto nuevo” (RON), “preferencia de objeto nuevo”, “preferencia espontánea del objeto” o “reconocimiento espontáneo del objeto”. Aquí haremos referencia a ella como la tarea RON.
Los descubrimientos recientes con las tareas VCP y RON son consistentes con los resultados que hacen uso de la tarea FIRM. Por ejemplo, los pacientes con deterioro de la memoria con daños que incluían el hipocampo mostraron deterioro de la memoria en la prueba VCP (McKee y Squire 1993). Además, los monos (Zola, et. al. 2000), las ratas (Clark, Zola y Squire 2000) y los ratones (Hammond, Tull y Stackman 2004) con daño selectivo del hipocampo, o con interrupción transitoria de la función del hipocampo, exhibieron alteraciones en la memoria dependiente de retraso cuando se probaba con la tarea VCP/RON (un resumen de los trabajos en animales con lesiones del hipocampo aparece en Squire, Wixted y Clark 2007; Clark y Martin 2005). La tarea RON ha sustituido en gran medida la tarea FIRM para la evaluación de la memoria de reconocimiento en animales de experimentación (Clark y Martin 2005; Winter, Saksida y Busset 2008).
LA ORGANIZACIÓN DE LA MEMORIA DECLARATIVA A LARGO PLAZO
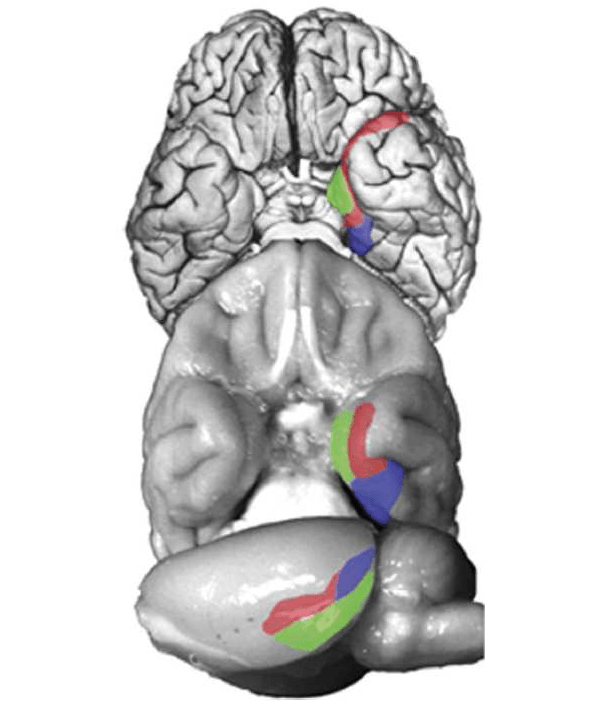
El trabajo en animales de experimentación durante las últimas tres décadas hizo uso de las anteriores tareas para identificar un sistema de estructuras anatómicamente conectadas en el lóbulo temporal medial que, cuando se daña, produce deterioro de la memoria como el primer deterioro revelado por el estudio de H.M. (véase Clark y Squire 2010). El sistema importante de las estructuras para la memoria incluye el hipocampo y las cortezas entorrinal, perirrinal, y parahipocampal adyacentes (Squire y Zola-Morgan 1991). La amígdala no es parte de este sistema. Se debe señalar también que en la rata, la corteza del hipocampo se conoce como la corteza postrinal, sin embargo, la organización anatómica es compartida en las tres especies (véase la Figura 1).
El hipocampo, que se define aquí como el GD, el cornu ammonis CA3 y el CA1, está situado anatómicamente para recibir información altamente procesada a partir de regiones generalizadas de la neocorteza a través de tres áreas corticales: las cortezas entorrinal, perirrinal, y parahipocampal, así como a través de otras proyecciones directas a la corteza entorrinal desde fuera del lóbulo temporal (véase la Figura 2). La vía principal para la transmisión de información sensorial en el hipocampo es la corteza entorrinal. La capa II de la corteza entorrinal proporciona su insumo principal. Esta proyección unidireccional, que forma parte de la vía perforante, proporciona un insumo sustancial al GD, que, a su vez, provee el principal insumo al CA3 a través de la proyección de fibras musgosas. También hay una proyección unidireccional más pequeña para el CA3 desde la capa II de la corteza entorrinal. El CA3 proporciona el principal insumo al CA1 vía la colateral de Schaffer/vía comisural, pero también hay una proyección asociativa recurrente sustancial dentro del campo de la CA3. El CA1 también recibe una proyección directa de la capa III de la corteza entorrinal, al igual que el subículo (la vía temporoamónica). Tanto las proyecciones de la colateral de Schaffer como las de la vía temporoamónica hacia el CA1 son unidireccionales. El CA1 proyecta principalmente hacia el subículo, pero también envía una proyección de la capa V de la corteza entorrinal. El subículo envía proyecciones principalmente a las capas IV y V de la corteza entorrinal (Witter y Amaral 2004).
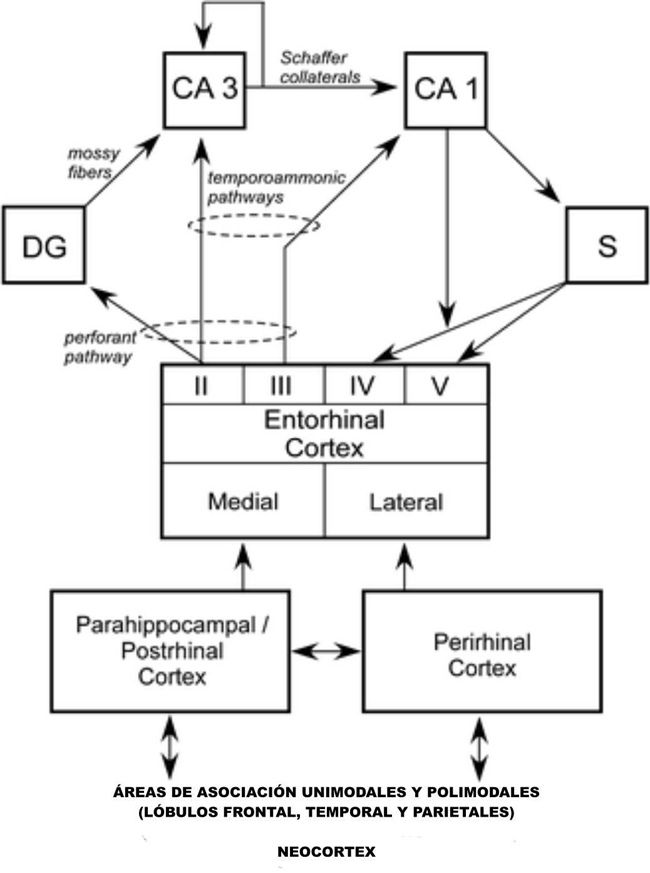
El hipocampo puede ser visto como aquello que reside al final del procesamiento jerárquico del lóbulo temporal medial, que recibe insumos tanto de las cortezas perirrinal y parahipocampal, así como de la corteza entorrinal (véase la Figura 2). Guiados por la anatomía, parece plausible que el hipocampo extiende y combina funciones ejecutadas por las estructuras que se proyectan a la misma (Squire, Wixted y Clark 2007). Hay que tener en cuenta también que las conexiones anatómicas de diferentes regiones de la neocorteza entran en el lóbulo temporal medial en diferentes puntos. Así, las áreas visuales más altas TE y TEO se proyectan preferentemente a la corteza perirrinal. A la inversa, la información espacial que va al lóbulo temporal medio de la corteza parietal llega exclusivamente a la corteza parahipocampal. De acuerdo con estos hechos anatómicos, se encontró que el daño en la corteza parahipocampal afecta la memoria espacial más que el daño en la corteza perirrinal (Parkinson, Murray y Mishkin 1988; Malkova y Mishkin 2003) y el daño en la corteza perirrinal afecta la ejecución de la tarea visual FIRM más que el daño en la corteza parahipocampal (Zola-Morgan, Squire y Rasmus 1994). Por último, la segregación relativa de la información visual y espacial parece mantenerse dentro de la corteza entorrinal, de manera que la corteza entorrinal lateral procesa más información visual y la corteza entorrinal medial procesa más información espacial.
EL RETO PARA EL MODELO ANIMAL Y LAS COMPARACIONES ENTRE ESPECIES
Como se analizó anteriormente, durante la década de 1960 y principios de 1970, el desarrollo de un modelo animal de la memoria humana y del deterioro de la memoria humana fue cuestionado por el hecho de que los animales podían hacer uso de la memoria no declarativa para resolver algunas tareas de memoria que los humanos normalmente abordan haciendo uso de la memoria declarativa. Por lo tanto, se volvió importante tanto entender bajo qué condiciones ocurre lo anterior, como identificar qué tipo de memoria se utiliza en cada caso. Este problema puede ilustrarse considerando al aprendizaje de discriminación simultánea, una tarea estándar que ha sido usada para estudiar la memoria de los mamíferos por más de cincuenta años. En una versión común de la tarea, se presentan ocho pares de objetos lado a lado cinco veces cada día, un par a la vez en un orden mezclado, para un total de cuarenta ensayos. Uno de los objetos de cada par es siempre correcto y la selección del objeto correcto produce una recompensa. La posición derecha/izquierda del objeto correcto varía de forma aleatoria entre los ensayos. Los seres humanos aprenden fácilmente esta tarea, ejecutando en un nivel mejor que el 80 por ciento de respuestas correctas después de sólo uno o dos días de entrenamiento (Squire, Zola-Morgan y Chen 1988). El que la ejecución dependa normalmente de la memoria declarativa, y de la memorización del objeto correcto en cada par, se demuestra por el hecho de que la ejecución de la tarea está altamente correlacionada con la capacidad para describir los objetos y por el hecho de que los pacientes con deterioro en la memoria tienen un desempeño pobre durante el periodo en que sus controles dominan la tarea. En contraste, esta misma tarea se adquirió a un ritmo normal en monos con lesiones en el hipocampo (Teng, Stefanacci, Squire y Zola 2000). Es importante destacar que los monos con daño en el hipocampo que también incluyen daños en el núcleo caudado (Teng, Stefanacci, Squite y Zola 2000) o los monos con daño selectivo al núcleo caudado (Fernandez-Ruiz, Wang, Aigner y Mishkin 2001) tuvieron un deterioro severo en el aprendizaje de la tarea. Estos descubrimientos sugieren que los monos aprenden la tarea de discriminación simultánea a través del ensayo/error, una estrategia de estímulo/respuesta que se basa en la memoria no declarativa. Si esto es así, surge naturalmente la pregunta de si los seres humanos también tienen una capacidad similar para la memoria no declarativa que podría apoyar el aprendizaje de esta misma tarea.

Para responder esta pregunta, a los pacientes con lesiones significativas en el lóbulo temporal medial, y esencialmente ninguna memoria declarativa residual, se les dio entrenamiento amplio en la tarea de discriminación simultánea durante un periodo de varias semanas (Bayley, Frascino y Squire 2005). La Figura 3A muestra la ejecución de los 4 participantes del grupo de control en la tarea. Estos participantes aprendieron la tarea con facilidad dentro de 3 sesiones. Un rasgo clave de la memoria declarativa es que puede ser expresada flexiblemente. Para demostrar este rasgo, se les dio a los individuos del grupo de control una tarea de clasificación de 3 a 6 días después del aprendizaje. Los 16 objetos usados en esta tarea se colocaron juntos en una mesa y se les dijo a los participantes que algunos de los objetos habían sido consistentemente diseñados como correctos. Se les pidió a continuación, para ordenar los objetos, colocar los objetos correctos de un lado de la mesa y los otros objetos en el otro lado. La Figura 3A muestra que los individuos del grupo de control no tenían dificultad con esta modificación de la tarea.
Por el contrario, el paciente E. P. aprendió gradualmente los pares de objetos en 36 sesiones de prueba (figura 3B), pero luego falló completamente en la tarea de clasificación cinco días después. Sin embargo, inmediatamente después, E.P. tuvo una buena ejecución en el formato estándar de la tarea (figura 3B). Diecisiete días más tarde, E. P. fracasó de nuevo en la tarea de clasificación, pero tuvo un buen desempeño en el formato original de la tarea. Los resultados para el paciente G.P. fueron esencialmente los mismos que para E. P. (figura 3C).
Algunas observaciones informales son reveladoras. Al comenzar cada día de prueba se les preguntó a los pacientes sobre los objetos y sobre lo que había ocurrido en las visitas anteriores. Sin embargo, nunca fueron capaces de recordar nada acerca de las sesiones de pruebas anteriores y no podían describir cómo habían usado los objetos. No obstante, como el entrenamiento avanzaba día a día, ambos pacientes (sin instrucciones explícitas sobre qué hacer en cada ensayo) tomaron fácilmente uno de los objetos de cada par y lo voltearon para recibir retroalimentación (la palabra “correcta” o “incorrecta” fue impresa debajo de la base de cada objeto).
Así, ambos pacientes adquirieron gradualmente ocho pares de objetos de la tarea de discriminación estándar durante muchas semanas; sin embargo, al inicio de cada sesión, no podían describir la tarea, las instrucciones o los objetos. El conocimiento adquirido era organizado de forma rígida y la ejecución colapsaba cuando el formato de la tarea se alteraba. Los resultados indican que los humanos poseen una capacidad sólida para el aprendizaje de ensayo y error gradual, que puede operar fuera de la conciencia de lo que se aprende y ser independiente del lóbulo temporal medial. Lo que se adquiere es inflexible y completamente diferente de la memoria declarativa. Estos resultados muestran que los humanos poseen un sistema de aprendizaje no declarativo como otros primates y sugieren cuáles son las características probables de este sistema (por ejemplo, que es inflexible e inaccesible a la conciencia).
En resumen, el desarrollo de un modelo de la memoria humana y su deterioro en el mono y la rata es una historia de éxito; el trabajo ha proporcionado una nueva percepción en la anatomía y la organización de la memoria de los mamíferos. Los retos pueden surgir debido a que estas especies han evolucionado bajo diferentes presiones selectivas que pueden sesgarlas a enfocar las tareas con estrategias diferentes a las usadas por los humanos. Al final, el análisis de estos casos, basado en nuestra comprensión de que hay múltiples sistemas de memoria con diferentes propiedades, ha servido para enfatizar las similitudes en la función de la memoria en las especies de mamíferos.
AGRADECIMIENTOS.
Este trabajo fue apoyado por el Servicio de Investigación Médica del Departamento de Asuntos de Veteranos, la beca 1120395 de la Fundación Nacional para la Ciencia y la beca 246000 del Instituto Nacional de Salud Mental. Traducción: Jonatan García Campos.

a) Centro Médico de Asuntos de Veteranos, San Diego, California, 92161, y los Departamentos de b) Psiquiatría, c) Neurociencias y d) Psicología de la Universidad de California en San Diego, La Jolla, California, 92093. / reclark@ucsd.edu
Ludus Vitalis, vol. XXI, num. 40, 2013, pp. 7997.
BIBLIOGRAFÍA
Bachevalier, J., Brickson, M., Hagger, C. (1993), “Limbicdependent recognition memory in monkeys develops early in infancy”, Neuroreport 4(1): 77–80.
Bayley, P.J., Frascino, J.C., Squire, L.R. (2005), “Robust habit learning in the absence of awareness and independent of the medial temporal lobe”, Nature 436(7050): 550–553.
Clark, R.E., Martin, S.J. (2005), “Interrogating rodents regarding their object and spatial memory” Curr Opin Neurobiol 15(5): 593–598.
Clark, R.E., Squire, LR. (2010), “An animal model of recognition memory and medial temporal lobe amnesia: History and current issues”, Neuropsychologia 48(8): 2234–2244.
Clark, R.E., Zola, S.M., Squire, L.R. (2000), “Impaired recognition memory in rats after damage to the hippocampus”, J Neurosci 20(23): 8853–8860.
Clark, R.E., West, A.N., Zola, S.M., Squire, L.R. (2001), “Rats with lesions of the hippocampus are impaired on the delayed nonmatchingtosample task”, Hippocampus 11(2): 176–186. 31.
Cohen, N.J., Squire, L.R. (1980), “Preserved learning and retention of patternanalyzing skill in amnesia: Dissociation of knowing how and knowing that”, Science 210(4466): 207–210.
Corkin, S., Amaral, D.G., González, RG., Johnson, K.A., Hyman, BT. (1997), “H. M.’s medial temporal lobe lesion: Findings from magnetic resonance imaging”, J Neurosci 17(10): 3964–3979.
Correll, R.E., Scoville, W.B. (1965), “Effects of medial temporal lesions on visual discrimination performance”, J Comp Physiol Psychol 60(2): 175–181.
Darwin, Charles (1859), On the Origin of Species. London: John Murray.
Desmond, Adrian (1984), Archetypes and Ancestors. Chicago: Univ of Chicago Press.
Douglas, R.J. (1967), “The hippocampus and behavior”, Psychol Bull 67(6): 416–422.
Ennaceur, A., Delacour, J. (1988), “A new onetrial test for neurobiological studies
of memory in rats. 1: Behavioral data”, Behav Brain Res 31(1): 47–59.
Fagan, J.F. (1970), “Memory in the infant”, J Exp Child Psychol 9(2): 217–226.
FernandezRuiz, J., Wang, J., Aigner, TG., Mishkin, M. (2001), “Visual habit formation in monkeys with neurotoxic lesions of the ventrocaudal neostriatum”, Proc Natl Acad Sci USA 98(7): 4196–4201.
Gaffan, D. (1974), “Recognition impaired and association intact in the memory of monkeys after transaction of the fornix”, J Comp Physiol Psychol 88(6): 1100–1109.
Gross, C.G. (1993a), “Hippocampus minor and man’s place in nature: A case study in the social construction of neuroanatomy”, Hippocampus 3(4): 403–415.
Gross, C.G. (1993b), “Huxley versus Owen: The hippocampus minor and evolution” ,Trends Neurosci 16(12): 493–498.
Hammond, R.S., Tull, LE., Stackman, RW. (2004), “On the delaydependent involvement of the hippocampus in object recognition memory”, Neurobiol Learn Mem 82(1): 26–34.
Huxley, Leonard (1900), Life and Letters of Thomas Henry Huxley. London: Macmillan. Huxley, T.H. (1861a), “On the zoological relations of man with the lower animals”, Natural History Review 1: 67–84.
Huxley, T.H. (1861b), “On the brain of Ateles paniscus”, Proceeding of the Zoological Society 247–260.
Huxley, Thomas Henry (1863), Evidence as to Man’s Place in Nature. London: Macmillan.
Kimble, D.P. (1963), “The effects of bilateral hippocampal lesions in rats”, J Comp Physiol Psychol 56: 273–283.
Lyell, Charles (1863), Geological Evidence of the Antiquity of Man. London: Spottiswoode & Co.
Malkova, L., Mishkin, M. (2003), “Onetrial memory for objectplace associations after separate lesions of hippocampus and posterior parahippocampal region in the monkey”, J Neurosci 23(5): 1956–1965.
McKee, R.D., Squire, LR. (1993), “On the development of declarative memory” J Exp Psychol Learn Mem Cogn 19(2): 397–404.
Milner, B. (1962), “Physiologie de l’Hippocampe”, ed. Passouant P (CNRS, Paris), pp. 257–272.
Mishkin, M. (1978), “Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus”, Nature 273(5660): 297–298.
Mishkin, M., Delacour, J. (1975), “An analysis of shortterm visual memory in the monkey”, J Exp Psychol Anim Behav Process 1(4): 326–334.
Mumby, D.G., Pinel, JP. (1994), “Rhinal cortex lesions and object recognition in rats”, Behav Neurosci 108(1): 11–18.
Mumby, D.G., Pinel, JP., Wood, ER. (1990), “Nonrecurringitems delayed nonmatchingtosample in rats: A new paradigm for testing nonspatial working memory”, Psychobiology 18: 321–326.
Owen, Richard (1859), On the Classification and Geographical Distribution of the Mammalia. London: Parker.
Parkinson, J.K., Murray, EA., Mishkin, MA. (1988), “A selective mnemonic role for the hippocampus in monkeys: Memory for the location of objects”, J Neurosci 8(11): 41594167.
Pavlov, Ivan (1927), Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex. trad. Anrep G.V., New York: Oxford Univ Press.
Ringo, J.L. (1993), “Spared shortterm memory in monkeys following medial temporal lobe lesions is not yet established: A reply to AlvarezRoyo, ZolaMorgan and Squire”, Behav Brain Res 59(12): 65–72.
Scoville, W.B., Milner, B. (1957), “Loss of recent memory after bilateral hippocampal lesions”, J Neurol Neurosurg Psychiatry 20(1): 11–21.
Squire, LR. (2004), “Memory systems of the brain: A brief history and current perspective”, Neurobiol Learn Mem 82(3): 171–177.
Squire, L.R. (2009), “The legacy of patient H.M. for neuroscience”, Neuron 61(1): 6–9.
Squire, L.R., ZolaMorgan, S. (1991), “The medial temporal lobe memory system”, Science 253(5026): 1380–1386.
Squire, L.R., Wixted, JT., Clark, R.E. (2007), “Recognition memory and the medial temporal lobe: A new perspective”, Nat Rev Neurosci 8(11): 872–883.
Squire, L.R., ZolaMorgan, S., Chen, K.S. (1988), “Human amnesia and animal models of amnesia: Performance of amnesic patients on tests designed for the monkey”, Behav Neurosci 102(2): 210–221.
Tang, Y.P., et al. (1999), “Genetic enhancement of learning and memory in mice”, Nature 401(6748): 63–69.
Teng, E., Stefanacci, L., Squire, L.R., Zola, S.M. (2000), “Contrasting effects on discrimination learning after hippocampal lesions and conjoint hippocampalcaudate lesions in monkeys”, J Neurosci 20(10): 3853–3863.
Weinstein, B. (1941), “Matchingfromsample by rhesus monkeys and by children”, J Comp Psychol 31: 195–213.
Winters, B.D., Saksida, L.M., Bussey, TJ. (2008), “Object recognition memory: Neurobiological mechanisms of encoding, consolidation and retrieval”, Neurosci Biobehav Rev 32(5): 1055–1070.
Witter, M.P., Amaral, D.G. (2004), “Hippocampal formation”, in G. Paxinos (ed.), The Rat Nervous System. San Diego: Elsevier Academic Press, pp. 637–703.
Zola, S.M., et al. (2000), “Impaired recognition memory in monkeys after damage limited to the hippocampal region”, J Neurosci 20(1): 451–463.
ZolaMorgan, S., Squire, L.R., Mishkin, M. (1982), “The neuroanatomy of amnesia: Amygdalahippocampus versus temporal stem”, Science 218(4579): 1337– 1339.
ZolaMorgan, S., Squire, L.R., Ramus, S.J. (1994), “Severity of memory impairment in monkeys as a function of locus and extent of damage within the medial temporal lobe memory system”, Hippocampus 4(4): 483–495.